
, L-alanil-L-glutamīna šķīdums")
Tilpums: 100 ml
Uzglabāšana: ≤ –15 °C
Sterilitāte: sterils filtrēts
Stabilais glutamīna šķīdums (L-alanil-L-glutamīns, 200 mM) ir ļoti stabila L-glutamīna dipeptīda forma, kas izstrādāta kā tiešs aizvietotājs parastajam L-glutamīnam šūnu kultūras barotnēs. L-glutamīns ir būtiska aminoskābe un galvenais enerģijas avots kultivētām šūnām, kas spēlē izšķirošu lomu šūnu augšanā, vielmaiņā un proteīnu sintēzē.
Pielietojums un priekšrocības
Standarta šķidrajā vidē L-glutamīns 37 °C temperatūrā salīdzinoši ātri sadalās, veidojot toksiskus blakusproduktus, piemēram, amonija jonus, kas var negatīvi ietekmēt šūnu dzīvotspēju un eksperimentu rezultātus. Stabilais glutamīns pārvar šo ierobežojumu, nodrošinot nesadalāmu dipeptīda formu, kas kultivēšanas apstākļos paliek neskarta.
Šūnas enzimātiski sadala dipeptīda saiti, lai atbrīvotu L-glutamīnu pēc nepieciešamības, nodrošinot nepārtrauktu svaigu piegādi un vienlaikus novēršot kaitīgu atkritumu produktu uzkrāšanos. Tas padara šo risinājumu īpaši izdevīgu ilgtermiņa šūnu kultūrām un augstas blīvuma augšanas sistēmām.
Sastāvs un lietošana
L-alanil-L-glutamīna šķīdums tiek pagatavots šūnu kultūras kvalitātes ūdenī 200 mM koncentrācijā un tiek sterili filtrēts, lai novērstu piesārņojumu jutīgos lietojumos. To var atšķaidīt tieši pilnīgā barotnē atbilstoši eksperimenta prasībām. Uzglabāt ≤ –15 °C temperatūrā un izvairīties no atkārtotas sasaldēšanas un atkausēšanas, lai saglabātu produkta stabilitāti.
Tikai pētniecības nolūkos. Nelietot diagnostikas vai terapeitiskās procedūrās. Nelietot cilvēkiem vai dzīvniekiem.
")
Tilpums: 100 ml
Uzglabāšana: +2 °C līdz +8 °C
Sterilitāte: sterils filtrēts
HEPES buferšķīdums (1 M), pazīstams arī kā N-2-hidroksietilpiperazīna-N-2-etānsulfoskābe, ir zwitterjoniskais organiskais buferšķīdums, ko plaši izmanto šūnu kultūras barotnēs. Tas ir paredzēts, lai uzturētu stabilu pH līmeni fizioloģiskajā diapazonā no 6,7 līdz 8,6, nodrošinot optimālu šūnu funkciju in vitro lietojumos.
Pielietojums un priekšrocības
HEPES nodrošina uzticamu buferēšanas spēju šūnu kultūras sistēmās, jo īpaši, ja šūnas tiek apstrādātas ārpus CO₂ inkubatora. 10–25 mM HEPES pievienošana kultūras barotnei nodrošina uzlabotu pH stabilitāti ilgstošas manipulācijas laikā, palīdzot uzturēt nemainīgus eksperimenta apstākļus.
Šis buferis ir necaurlaidīgs membrānām, minimāli traucē bioķīmiskās reakcijas un demonstrē spēcīgu ķīmisko un enzīmatisko stabilitāti. Šīs īpašības padara to piemērotu plašam šūnu kultūru un bioķīmisko lietojumu klāstam.
Sastāvs un lietošana
Šķīdums tiek piegādāts kā 1 M koncentrāts, kas sagatavots šūnu kultūras kvalitātes ūdenī, un ir sterili filtrēts lietošanai vidē, kas ir jutīga pret piesārņojumu. To var atšķaidīt līdz vēlamajai darba koncentrācijai atkarībā no lietošanas prasībām. Uzglabāt +2 °C līdz +8 °C temperatūrā un apstrādāt aseptiskos apstākļos, lai saglabātu produkta integritāti.
Tikai pētniecības nolūkos. Nelietot diagnostikas vai terapeitiskās procedūrās. Nelietot cilvēkiem vai dzīvniekiem.
, 250 g/l")
Tilpums: 50 ml
Uzglabāšana: +2 °C līdz +8 °C
Sterilitāte: sterils filtrēts
D-(+)-glikozes šķīdums (dekstrozes šķīdums) ir sterils, lietošanai gatavs uztura bagātinātājs, kas satur dabīgi sastopamo cukuru D-(+)-glikozi, kas ir galvenā šūnu vielmaiņas sastāvdaļa. Glikoze piedalās būtiskos bioloģiskos procesos, piemēram, enerģijas ražošanā, glikozilācijā un glikānu veidošanā, kas veicina šūnu struktūru un funkcijas.
Lietošana un priekšrocības
Šis glikozes šķīdums tiek plaši izmantots kā piedeva šūnu kultūras barotnēs un daudzās šūnu un molekulārās bioloģijas lietojumprogrammās. Kā primārais oglekļa un enerģijas avots glikoze atbalsta šūnu augšanu, proliferāciju un metabolisko aktivitāti. Tās līdzdalība biosintēzes ceļos padara to ļoti svarīgu normālas šūnu fizioloģijas un eksperimentu konsekvences uzturēšanai.
Sastāvs un lietošana
Šķīdums tiek piegādāts ar augstu glikozes koncentrāciju (250 g/l), kas ļauj to elastīgi atšķaidīt kultūras barotnēs atbilstoši eksperimenta prasībām. Tas ir sterili filtrēts, lai nodrošinātu piemērotību lietojumiem, kas ir jutīgi pret piesārņojumu. Uzglabāt +2 °C līdz +8 °C temperatūrā un rīkoties aseptiski, lai saglabātu produkta kvalitāti un stabilitāti.
Tikai pētniecības nolūkos. Nelietot diagnostikas vai terapeitiskās procedūrās. Nelietot cilvēkiem vai dzīvniekiem.
, šķīdums, 100x")
Tilpums: 10 ml
Uzglabāšana: +2 °C līdz +8 °C
Sterilitāte: Sterili filtrēts
Insulīna-transferīna-selēna (ITS) šķīdums (100x) ir ķīmiski definēts papildinājums, kas paredzēts plašam šūnu kultūru pielietojumu klāstam. To visbiežāk izmanto kā piedevu bazālajām šūnu kultūru barotnēm, lai atbalstītu šūnu augšanu samazināta seruma vai bezseruma apstākļos.
Lietošana un priekšrocības
Mūsu ITS piedeva nodrošina būtiskas sastāvdaļas, kas nepieciešamas bezseruma barotņu optimālai darbībai. Papildinot tradicionālās barotnes ar ITS, var ievērojami samazināt nepieciešamību pēc teļa seruma (FBS) daudzu šūnu līniju ikdienas uzturēšanai. Tas palīdz samazināt ar seruma lietošanu saistīto variabilitāti, vienlaikus saglabājot stabilu šūnu augšanu un dzīvotspēju.
Insulīns atbalsta galveno uzturvielu uzsūkšanos šūnās un vielmaiņu, transferīns veicina dzelzs transportu, bet selēns veicina antioksidantu aizsardzību un fermentu aktivitāti. Kopā šīs sastāvdaļas veicina līdzsvarotu šūnu vielmaiņu un uzlabo reproducējamību noteiktās kultivēšanas sistēmās.
Sastāvs un lietošana
Insulīns-transferīns-selēns (ITS) tiek piegādāts kā 100x koncentrēts šķīdums Earle's Balanced Salt Solution (EBSS) bez fenola sarkana. Standarta lietošanai atšķaidiet 1:100 attiecīgajā pamatbarotnē, lai sasniegtu ieteicamo darba koncentrāciju. Uzglabājiet +2 °C līdz +8 °C temperatūrā un rīkojieties aseptiskos apstākļos, lai saglabātu produkta stabilitāti un sterilitāti.
Tikai pētniecības vajadzībām. Nelietot diagnostiskās vai terapeitiskās procedūrās. Nelietot cilvēkiem vai dzīvniekiem.
")
Tilpums: 5 ml
Uzglabāšana: +2 °C līdz +8 °C
Sterilitāte: Sterili filtrēts
Recombinant Human Insulin Solution ir ķīmiski definēts papildinājums, ko parasti izmanto zīdītāju šūnu līniju, tostarp Ķīnas kāmju olnīcu (CHO) šūnu, kultivēšanai. Šis šūnu kultūras kvalitātes šķīdums satur rekombinantā cilvēka insulīnu, kas ekspresēts Saccharomyces cerevisiae, nodrošinot augstu tīrību un stabilu darbību pētniecības lietojumos.
Lietošana un priekšrocības
Insulīnu parasti pievieno bezseruma un ķīmiski definētiem barotnēm, lai veicinātu šūnu augšanu un produktivitāti. Kā galvenais regulējošais hormons insulīns atbalsta glikozes, aminoskābju un taukskābju uzsūkšanos, izmantošanu un uzkrāšanu šūnās. Tas arī kavē glikogēna, proteīnu un lipīdu noārdīšanos, tādējādi veicinot uzlabotu šūnu dzīvotspēju un metabolisko stabilitāti kultivēšanas sistēmās. Ķīmiski definētā formula nodrošina reproducējamību un samazina variabilitāti jutīgās šūnu kultivēšanas darba plūsmās.
Bioloģiskās īpašības un lietošana
Insulīns ir divķēžu polipeptīda hormons, ko dabīgi ražo aizkuņģa dziedzera salu β-šūnas. Tā molekulmasa ir aptuveni 5800 Da. α
- un β-ķēdes ir savienotas ar divām starpķēžu disulfīda saitēm, un α-ķēdē ir viena iekšķēžu disulfīda saite. Šūnu kultūru lietojumiem šķīdums jāapstrādā aseptiskos apstākļos un jāuzglabā temperatūrā no +2 °C līdz +8 °C, lai saglabātu stabilitāti un efektivitāti.
Tikai pētniecības vajadzībām. Nelietot diagnostiskās vai terapeitiskās procedūrās. Nelietot cilvēkiem vai dzīvniekiem.
")
Tilpums: 100 ml
Uzglabāšana: +2 °C līdz +8 °C
Sterilitāte: sterils filtrēts
Nātrija piruvāta šķīdums (100 mM) ir sterils, lietošanai gatavs papildinājums, kas paredzēts, lai nodrošinātu papildu, viegli pieejamu enerģijas avotu šūnu kultūras barotnēm. Nātrija piruvāts spēlē galveno lomu šūnu enerģijas vielmaiņā un atbalsta metaboliski aktīvu un strauji vairojošos šūnu, piemēram, audzēju šūnu, augšanu. Papildinājums var uzlabot šūnu dzīvotspēju un palīdzēt uzturēt metabolisko stabilitāti kultūras sistēmās.
Pielietojums un priekšrocības
Šis šķīdums tiek plaši izmantots ikdienas šūnu kultivēšanā, lai bagātinātu barotni ar piruvātu un veicinātu optimālus augšanas apstākļus. Tas atbalsta ATP ražošanu, var palīdzēt samazināt oksidatīvo stresu un veicina kultivēto šūnu vielmaiņas uzlabošanos. Ražots šūnu kultivēšanai piemērotā ūdenī un sterilizēts filtrējot, produkts nodrošina nemainīgu kvalitāti un reproducējamību pētniecības darba procesā.
Lietošana un saderība
Ieteicamā galīgā koncentrācija lielākajai daļai šūnu kultivēšanas lietojumu ir 1 mM nātrija piruvāts, ko iegūst, atšķaidot 100 mM pamatšķīdumu 1:100 pilnīgā kultivēšanas vidē. Šķīdums ir saderīgs ar plašu bāzes barotņu un zīdītāju šūnu līniju klāstu. Uzglabāt +2 °C līdz +8 °C temperatūrā un aizsargāt no piesārņojuma, lai saglabātu produkta stabilitāti.
Tikai pētniecības nolūkos. Nelietot diagnostikas vai terapeitiskās procedūrās. Nelietot cilvēkiem vai dzīvniekiem.
")
Tilpums: 100 ml Uzglabāšana: ≤-15°C Sterilitāte: Sterilitāte: Sterili jāfiltrē
Antibiotiku/antimikotisko līdzekļu šķīdums (100x ) ir sterils, lietošanai gatavs koncentrāts, kas paredzēts mikrobioloģiskā piesārņojuma risku samazināšanai šūnu kultūru un saistītās laboratorijās. Šis 100x šķīdums satur vispāratzītu penicilīna, streptomicīna un amfotericīna B kombināciju, kas nodrošina plaša spektra antimikrobiālu iedarbību pret grampozitīvām un gramnegatīvām baktērijām, raugiem un pavedienveida sēnītēm. Preparāts ir piemērots lietošanai eikariotisko šūnu kultūrās, bakteriālajās barotnēs un citās sistēmās, kas jutīgas pret piesārņojumu, veicinot tīras un konsekventas laboratorijas darbības.
Lietošana un priekšrocības Šis šķīdums, kas optimizēts ikdienas pētniecības protokoliem, tiek plaši izmantots, lai uzturētu aseptiskus apstākļus šūnu kultūru darba plūsmās. Tas nodrošina uzticamu veiktspēju vidē, kas jutīga pret piesārņojumu, palīdzot pētniekiem samazināt mikrobu pārmērīgas augšanas risku, neapdraudot šūnu veselību vai eksperimentālo reproducējamību. Sterilā, filtrētā formula novērš nepieciešamību pēc papildu šķīdināšanas posmiem, veicinot racionalizētu barotņu sagatavošanu un samazinot mainīgumu ikdienas laboratorijas procedūrās.
Lietošana un saderība Lai sasniegtu standarta darba koncentrācijas, atšķaidiet šķīdumu 1:100 pilnā barotnē. Produkts ir saderīgs ar plašu zīdītāju šūnu līniju un pamatnes barotņu klāstu. Pastāvīga krājumu pieejamība ļauj pētniekiem gūt labumu no uzticamas piegādes nepārtrauktības un vienkāršotas loģistikas plānošanas. Lai saglabātu šķīduma stabilitāti, tas jāuzglabā ≤ -15 °C temperatūrā un jāaizsargā no atkārtotiem sasalšanas un atkausēšanas cikliem. Tikai izmantošanai pētniecībā. Nav paredzēts lietošanai diagnostikas vai terapeitiskās procedūrās. Nav paredzēts lietošanai cilvēkiem vai dzīvniekiem.
")
Tilpums: 100 ml Uzglabāšana: + 2°C līdz +8°C Sterilitāte: Sterilitāte: Sterili filtrēts
MEM Non-Essential Amino Acids (100x ) ir sterils papildinājums, kas paredzēts šūnu augšanas un dzīvotspējas uzlabošanai zīdītāju šūnu kultūru sistēmās. Sastāvs atbilst 100x koncentrētam neaizvietojamo aminoskābju koncentrātam, kas atrodamas standarta minimālajā pamatvielā (MEM), ļaujot ar minimālu sagatavošanas darbu tieši papildināt bāzes barotnes.
Pielietojums un priekšrocības Šis papildinājums nodrošina papildu aminoskābju rezervi strauji proliferējošām šūnām vai šūnu līnijām, kas ir zaudējušas spēju sintezēt neaizvietojamās aminoskābes de novo. Samazinot biosintēzes radīto metabolisko slogu, tas veicina uzlabotu augšanas kinētiku, ilgāku dzīvotspēju un lielāku eksperimentālo konsekvenci
- īpaši attiecībā uz jutīgām pret barības vielām vai augsta blīvuma kultūrām.
Sastāvs un lietošana Šķīduma sastāvā ir glicīns, L-alanīns, L-asparagīns, L-asparagīnskābe, L-glutamīnskābe, L-prolīns un L-serīns. Tas ir saderīgs ar MEM un lielāko daļu citu standarta barotņu. Lai izmantotu, atšķaidiet 1:100 galīgajā barotnē. Šis produkts ir sterili filtrēts un gatavs lietošanai bez papildu apstrādes darbībām. Tikai izmantošanai pētniecībā. Nav paredzēts lietošanai diagnostikas vai terapeitiskās procedūrās. Nav paredzēts lietošanai cilvēkiem vai dzīvniekiem.

Accutase ir gatavs lietošanai, sterili filtrēts šūnu atdalīšanas šķīdums, kas izstrādāts kā maiga alternatīva tripsīnam/EDTA, lai atdalītu piekļāvušās zīdītāju šūnas no standarta audu kultūras plastmasas traukiem un ar adhezīvu pārklātām virsmām. Tas apvieno proteolītisko un kolagenolītisko enzīmu aktivitāti sabalansētā sāls šķīdumā, lai nodrošinātu efektīvu, bet kontrolētu atdalīšanu, saglabājot šūnu virsmas proteīnus un veicinot augstu dzīvotspēju pēc pārneses un ātru atkārtotu piekļāvēšanos.
Accutase sastāvs ir balstīts uz Dulbecco fosfāta buferšķīdumu (DPBS) ar EDTA un fenolsarkano kā vizuālu pH indikatoru. Fermenti ir ne-zīdītāju un ne-baktēriju izcelsmes, padarot Accutase īpaši piemērotu cilmes šūnu pētniecībai, vakcīnu ražošanas procesiem un jebkurai lietošanai, kurā ir jāminimizē dzīvnieku vai mikrobiālas izcelsmes piesārņotāji. Šķīdums automātiski inhibējas 37 °C temperatūrā, tādēļ pēc atdalīšanas nav nepieciešams neitralizējošs reaģents vai serumu saturošs barotne – šūnas var pārnest tieši svaigā barotnē.
Galvenās īpašības
Lietošanai gatavs 1x sterili filtrēts šķidrums – nav nepieciešama atšķaidīšana vai atjaunošana
Kombinēta proteolītiskā un kolagenolītiskā enzīmu aktivitāte maigai disociācijai
Katra partija standartizēta atbilstoši noteiktam disociācijas aktivitātes līmenim, lai nodrošinātu partiju savstarpēju atbilstību
Enzīmu izcelsme nav saistīta ar zīdītājiem vai baktērijām
Automātiska inhibīcija 37 °C temperatūrā – nav nepieciešams neitralizējošs šķīdums
Formulēts Dulbecco PBS ar EDTA
Fenolsarkans iekļauts kā vizuāls pH indikators
pH 6,8–7,8
Tipiski pielietojumi
Accutase maigi disociē plašu klāstu adhezīvu un jutīgu šūnu tipu, tostarp cilvēka embrionālās cilmes šūnas (hESCs), cilvēka indukcijas pluripotentās cilmes šūnas (iPSCs), neironu cilmes šūnas, primāros neironus un rutīniski kultivētas adhezīvās šūnu līnijas, piemēram, HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 un A549. Tipiski lietošanas gadījumi ietver:
Rutīniska adhezīvo zīdītāju šūnu subkultivēšana un pasāžēšana
Maiga hESCs, iPSCs un citu jutīgu līniju atsevišķu šūnu atdalīšana
Paraugu sagatavošana plūsmas citometrijai un FACS analīzei
Šūnu virsmas marķieru analīze, kur svarīga ir epitopa integritāte
Šūnu migrācijas, proliferācijas un apoptozes analīzes
Miera stāvokļa testi, izmantojot seruma izsīkšanu un onkogēnu transfekcijas pētījumus
Audzēja šūnu un neirālo krastu šūnu migrācijas testi
Ražošanas apjoma palielināšana bioreaktoru darba plūsmās
Ikdienas darbā uzklājiet aptuveni 10 ml Accutase uz 75 cm2 kultūras virsmas un inkubējiet 5–10 minūtes istabas temperatūrā. Optimālais inkubācijas laiks jānosaka katrai šūnu līnijai atsevišķi, un tas nedrīkst pārsniegt vienu stundu. Pirms pievienošanas noskalojiet šūnu slāni ar Ca2+/Mg2+-nesaturošu sāls šķīdumu, piemēram, DPBS bez kalcija un magnija, lai noņemtu seruma atliekas un divvērtīgos katjonus.
Rīkošanās un uzglabāšana
Neatvērtu pudeli uzglabājiet sasaldētu -15 °C vai zemākā temperatūrā. Atkausējiet istabas temperatūrā vai nakti +2 °C līdz +8 °C temperatūrā. Neatkausējiet Accutase 37 °C ūdens vannā, jo paaugstināta temperatūra samazina fermentu aktivitāti. Pēc atkausēšanas šķīdumu var uzglabāt līdz 2 mēnešiem +2 °C līdz +8 °C temperatūrā; neuzglabājiet istabas temperatūrā. Pirms lietošanas reaģentu neiesildiet līdz 37 °C – pievienojiet to tieši izskalotām šūnām istabas temperatūrā. Lai nodrošinātu ilgu glabāšanas laiku, ieteicams izmantot vienreizējās lietošanas alikvotas, lai izvairītos no atkārtotiem atkausēšanas cikliem. Vienmēr strādājiet aseptiskos apstākļos.
Kvalitāte
Ražots saskaņā ar stingriem kvalitātes standartiem. Katra Accutase partija tiek sterilizēta ar filtrēšanu un pārbaudīta attiecībā uz sterilitāti, pH, izskatu un disociācijas aktivitāti, lai nodrošinātu vienmērīgu, reproducējamu darbību no partijas uz partiju.
Produkta specifikācijas
Specifikācija
Sīkāka informācija
Produkta veidsŠūnu atdalīšanas / disociācijas reaģents
FormātsSterili filtrēts šķidrums, gatavs lietošanai
Tilpums100 ml
Darba koncentrācija1x (gatavs lietošanai)
Enzīmu aktivitāteKombinēta proteolītiska un kolagenolītiska
Fermentu izcelsmeNe no zīdītājiem un ne no baktērijām
Buferu sistēmaDulbecco PBS ar EDTA
pH indikatorsFenolsarkans
pH diapazons6,8 – 7,8
IzskatsDzidrs, gaiši sarkans līdz oranžs šķīdums
Uzglabāšanas temperatūra-15 °C vai zemāka
Stabilitāte pēc atkausēšanasLīdz 2 mēnešiem temperatūrā no +2 °C līdz +8 °C
Ieteicamais lietošanas daudzums~10 ml uz 75 cm2 kultūras virsmas
Tipisks inkubācijas laiks5–10 minūtes istabas temperatūrā
Piegādes nosacījumiSasaldēts uz sausā ledus
Paredzētais lietojumsTikai pētniecības vajadzībām un turpmākai ražošanai
Sastāvs (sastāvs uz litru)
Sastāvdaļa
Koncentrācija (mg/l)
Neorganiskie sāļi
Nātrija hlorīds (NaCl)8000,00
Dinātrija hidrogēnfosfāts (Na2HPO4)1150,00
Kālija hlorīds (KCl)200,00
Kālija dihidrogēnfosfāts (KH2PO4)200,00
Citas sastāvdaļas
EDTA · 4Na (tetranātrija EDTA)220,00
Fenolsarkans3,00
Patentēts fermentu maisījums (proteolītiska un kolagenolītiska aktivitāte)1x
Accutase ir Innovative Cell Technologies, Inc. reģistrēta preču zīme.
, w: 2 mM L-glutamīns, w: 2,2 g/L NaHCO3, w: EBSS")
Šis lietošanai gatavs, sterili filtrēts šķidrais preparāts ir papildināts ar Earle’s Balanced Salt Solution (EBSS), 2 mM L-glutamīnu, D-glikozi (1,0 g/l) un 2,2 g/l nātrija bikarbonātu (NaHCO3), tādējādi padarot to piemērotu lietošanai CO2 kontrolētā inkubatora vidē (parasti 5 % CO2). Iekļautais fenolsarkans darbojas kā pH indikators, ļaujot ērti vizuāli uzraudzīt barotnes stāvokli šūnu kultivēšanas laikā.
Galvenās īpašības
Klasiskais Eagle MEM sastāvs ar Earle’s Balanced Salt Solution (EBSS)
Iekļauts 2 mM L-glutamīns – gatavs tūlītējai lietošanai
2,2 g/l nātrija bikarbonāta – buferēts 5 % CO2 inkubācijai
Ar D-glikozi (1,0 g/l) kā galveno oglekļa avotu
Ar fenolsarkano kā pH indikatoru
Bez HEPES un bez nātrija piruvāta
Sterili filtrēta šķidrā barotne, gatava lietošanai
pH 7,0 – 7,6
Tipiski pielietojumi
EMEM atbalsta dažādu zīdītāju šūnu līniju kultivēšanu, tostarp HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 un daudzas primārās šūnas. Tipiski pielietojumi ietver:
Adhezīvo šūnu līniju ikdienas uzturēšana un pavairošana
Vīrusu pavairošana un vakcīnu ražošanas procesi
Citotoksicitātes un bioanalīžu lietojumi
Transfekcijas un proteīnu ekspresijas pētījumi
Pamatpētījumi šūnu bioloģijā un molekulārajā bioloģijā
Optimālai šūnu augšanai EMEM parasti papildina ar 5–10 % teļa seruma (FBS) un, atkarībā no šūnu līnijas, ar neesamajām aminoskābēm (NEAA) un antibiotikām, piemēram, penicilīnu/streptomicīnu.
Rīkošanās un uzglabāšana
Neatvērtu pudeli uzglabājiet +2 °C līdz +8 °C temperatūrā, aizsargājot no gaismas. Pēc atvēršanas lietojiet aseptiskos apstākļos. L-glutamīns šķīdumā pakāpeniski sadalās – lai nodrošinātu labāko rezultātu, ieteicams izmantot barotni 4 nedēļu laikā pēc atvēršanas vai, ja tā tiek uzglabāta ilgāku laiku, pirms lietošanas papildināt ar svaigu L-glutamīnu. Pirms pievienošanas šūnām ļaujiet barotnei iesilt līdz 37 °C.
Kvalitāte
Ražots saskaņā ar stingriem kvalitātes standartiem. Katra partija tiek pārbaudīta uz sterilitāti, pH, osmolalitāti un endotoksīnu līmeni, lai nodrošinātu stabilu darbību šūnu kultūru lietojumos.
Produkta specifikācijas
Specifikācija
Detalizēta
Produkta veidsMEM
Produkta kategorijaŠūnu kultūras barotnes
FormātsŠķidrums
SterilsJā
Izmērs500 ml
L-glutamīnsAr L-glutamīnu (2 mM)
GlikozesAr glikozes (1,0 g/l)
Nātrija bikarbonātsAr NaHCO3 (2,2 g/l)
HEPESBez HEPES
Nātrija piruvātsBez nātrija piruvāta
FenolsarkansAr fenola sarkano
Sāls šķīdumsEarle’s Balanced Salt Solution (EBSS)
pH7,0 – 7,6
Endotoksīnu satursNav norādīts
Uzglabāšana+2 °C līdz +8 °C
Sastāvs (sastāvs uz litru)
Sastāvdaļa
Koncentrācija (mg/l)
Neorganiskie sāļi
Kalcija hlorīds · 2H2O265,00
Magnija sulfāts97,72
Kālija hlorīds400,00
Nātrija hlorīds6 800,00
Nātrija dihidrogēnfosfāts, bezūdens122,00
Nātrija bikarbonāts (NaHCO3)2 200,00
Aminoskābes
L-arginīns · HCl126,00
L-cistīns · 2HCl31,30
L-glutamīns292,00
L-histidīns · HCl · H2O42,00
L-izoleicīns52,00
L-leicīns52,00
L-lizīns · HCl72,50
L-metionīns15,00
L-fenilalanīns32,00
L-treonīns48,00
L-triptofāns10,00
L-tirozīns · 2Na · 2H2O51,90
L-valīns46,00
Vitamīni
D-kalcija pantotenāts1,00
Holīna hlorīds1,00
Folijskābe1,00
mio-inozitols2,00
Nikotinamīds1,00
Piridoksāls · HCl1,00
Riboflavīns0,10
Tiamīns · HCl1,00
Citas sastāvdaļas
D(+)-glikoze1 000,00
Fenolsarkans10,00

Freeze Medium CM-1 galvenās īpašības:
Plaša saderība: Efektīva dažādiem šūnu tipiem, tostarp primārajām šūnām, cilmes šūnām un izveidotām šūnu līnijām.
Augsta dzīvotspēja: Optimizēta, lai pēc atkausēšanas maksimāli palielinātu šūnu reģenerāciju un dzīvotspēju, nodrošinot uzticamus eksperimentu rezultātus.
Gatavs lietošanai: Ērti sagatavotas un sterilizētas tūlītējai lietošanai, samazinot sagatavošanas laiku un piesārņojuma risku.
Uzlabota stabilitāte: Uztur nemainīgu veiktspēju standarta kriokonservēšanas apstākļos, nodrošinot reproducējamus rezultātus.
Ilgs glabāšanas laiks: CM-1 ir serumu saturoša, lietošanai gatava kriokonservēšanas barotne, ko var uzglabāt ledusskapī līdz pat vienam gadam.
CM-1 izmantošana šūnu sasaldēšanai
Lai izmantotu CM-1 gan adhēzijas, gan suspensijas šūnu sasaldēšanai, izpildiet šādas darbības
Attiecībā uz adhēzīvām šūnām tās nomazgā un atdala no kultūras substrāta. Suspensijas šūnām pārejiet uz nākamo soli.
Saskaitiet šūnas, lai pārliecinātos, ka to koncentrācija ir atbilstoša.
Centrifugējiet šūnas, lai tās granulētu, pēc tam atkārtoti suspendējiet CM-1 sasaldēšanas barotnē.
Resuspendētās šūnas pārvietot kriokivās.
Pirms šūnu pārvietošanas uz ilgtermiņa uzglabāšanu izmantojiet lēnas sasaldēšanas metodi
Metode
Apraksts
Soļi
❄️
Manuāla sasaldēšana
Pakāpeniska metode, kas ietver pakāpenisku temperatūras pazemināšanu, lai nodrošinātu šūnu dzīvotspēju
1️⃣ Ievietojiet šūnas sasaldēšanas barotnē 4 °C saldētavā uz 40 minūtēm.
2️⃣ Uz 24 stundām pārvietot uz -80°C saldētavu.
3️⃣ Ilgtermiņa uzglabāšanai šūnas uzglabāt šķidrā slāpeklī
❄️
Frosty kunga izmantošana
Ērta ierīce, kas ļauj kontrolēt sasaldēšanas ātrumu bez elektriskās strāvas
1️⃣ Sagatavojiet šūnas krioviālos ar sasaldēšanas barotni.
2️⃣ Ievietojiet krioviālus Mr. Frosty konteinerā.
3️⃣ Pirms pārvietošanas uz šķidro slāpekli 24 stundas uzglabāt -80 °C temperatūrā
❄️
Kontrolēta ātruma saldētava
Thermo Fisher vai citu ražotāju augstas precizitātes saldētava, kas paredzēta kontrolētai temperatūras pazemināšanai
1️⃣ Ierīci ieprogrammējiet tā, lai pakāpeniski pazeminātu temperatūru.
2️⃣ Ievietojiet sagatavotās šūnas saldētavā.
3️⃣ Pēc sasaldēšanas cikla šūnas pārceliet uz šķidro slāpekli
Glabājiet krioviālus temperatūrā, kas zemāka par -130 °C, vai šķidrā slāpeklī ilgstošai uzglabāšanai.
Sastāvdaļas
Satur FBS, DMSO, glikozi, glikozi, sāļus
Buferizācijas spēja: pH = 7,2 līdz 7,6
Cytion's Freeze Medium CM-1 piedāvā uzticamu risinājumu kriokonservēšanai, nodrošinot augstu šūnu dzīvotspēju un funkcionalitāti pēc atkausēšanas visdažādākajiem pētniecības lietojumiem.

- specializētu Ham's F-12 barotnes modifikāciju, kas izstrādāta, lai atbilstu unikālām bioloģisko pētījumu prasībām. Šī uzlabotā barotne piedāvā īpašas priekšrocības, uzlabojot primāro cilvēka hepatocītu, kā arī žurku un vistu aknu šūnu kultivēšanu, jo īpaši pazemināta seruma apstākļos.
Ham's F-12K (Kaighn's) barotne ir rūpīgi izstrādāta, lai optimizētu šūnu kultūras apstākļus. Tai ir bagātināts sastāvs, kas nodrošina paaugstinātu tādu būtisku sastāvdaļu kā aminoskābju un nātrija piruvāta, kā arī papildu elementu, tostarp putrescīna, timidīna, hipoksantīna un cinka, līmeni. Šie papildinājumi ļauj pētniekiem papildināt barotni ar minimālu seruma daudzumu vai noteiktiem komponentiem konkrētiem šūnu tipiem, tādējādi atvieglojot precīzus eksperimentālos apstākļus.
Hama F-12K (Kaigna) barotne nesatur olbaltumvielas vai augšanas faktorus. Tāpēc bieži vien ir nepieciešama papildināšana ar augšanas faktoriem un Fetal Bovine Serum (FBS), kas ļauj pētniekiem pielāgot barotni savu konkrēto šūnu līniju prasībām. Lai nodrošinātu optimālu veiktspēju, FBS koncentrācija ir rūpīgi jāoptimizē katrai šūnu līnijai, nodrošinot optimālu augšanu un funkcionalitāti.
Lai uzturētu fizioloģisko pH, Hama F-12K (Kaighn's) barotnē izmanto nātrija bikarbonāta buferisko sistēmu (2,5 g/l), tāpēc kultivēšanas laikā ir nepieciešama kontrolēta 5-10 % CO2 vide. Tas nodrošina, ka barotnes pH saglabājas ideālā diapazonā šūnu augšanai un dzīvotspējai
Kvalitātes kontrole
pH = 7,2 +/
- 0,02 20-25 °C temperatūrā.
Katra partija ir pārbaudīta attiecībā uz sterilitāti un mikoplazmas un baktēriju neesamību.
Uzturēšana
Glabāt ledusskapī +2°C līdz +8°C temperatūrā tumsā. Sasaldēšana, kā arī sasilšana līdz +37°C samazina produkta kvalitāti.
Nesildiet barotni līdz temperatūrai, kas pārsniedz 37°C, un neizmantojiet nekontrolējamus siltuma avotus (piemēram, mikroviļņu krāsnis).
Ja jāizmanto tikai daļa barotnes, izņemiet šo daudzumu no pudeles un uzsildiet to istabas temperatūrā.
Jebkuras barotnes, izņemot pamata barotni, derīguma termiņš ir 8 nedēļas no izgatavošanas datuma.
Sastāvs
Sastāvdaļas
mg/l
Neorganiskie sāļi
Kalcija hlorīds x 2H2O
135,24
Vara(II) sulfāts x 5H2O
0,00
Dzelzs (II) sulfāts x 7H2O
0,83
Magnija hlorīds x 6H2O
105,72
Magnija sulfāts x 7H2O
394,49
Kālija hlorīds
283,29
Kālija dihidrogēnfosfāts
58,52
Nātrija hlorīds
7597,20
nātrija hidrogēnfosfātsbezūdens
115,02
Cinka sulfāts x 7H2O
0,14
Citi komponenti
D(+)-glikoze, bezūdens viela
1260,00
Hipoksantīns
4,08
DL-α-liposkābe
0,21
Fenola sarkanais
3,00
Putrescīns x 2HCl
0,32
Nātrija piruvāts
220,00
NaHCO3
2500,00
Timidīns
0,73
Aminoskābes
L-alanīns
17,82
L-arginīns x HCl
421,40
L-asparagīns x H2O
30,02
L-asparagīnskābe
26,62
L-cisteīns x HCl x H2O
70,24
L-glutamīns
292,20
L-glutamīnskābe
29,42
Glicīns
15,01
L-histidīns x HCl x H2O
41,92
L-izoleicīns
7,87
L-leicīns
26,24
L-lizīns x HCl
73,04
L-metionīns
8,95
L-fenilalanīns
9,91
L-Prolīns
69,06
L-serīns
21,02
L-treonīns
23,82
L-triptofāns
4,08
L-tirozīns
10,87
L-valīns
23,42
Vitamīni
D(+)-biotīns
0,07
D-kalcija pantotenāts
0,48
Holīna hlorīds
13,96
Folijskābe
1,32
mio-inositols
18,02
Nikotīnamīds
0,04
Piridoksīns x HCl
0,06
Riboflavīns
0,04
Tiamīns x HCl
0,34
B12 vitamīns
1,36

Fosfātu buferšķīdums (PBS) ir plaši izmantots buferšķīdums bioloģiskajos un ķīmiskajos pētījumos. Tam ir izšķiroša nozīme pH līdzsvara un osmolaritātes uzturēšanā dažādu eksperimentālu procedūru laikā, tostarp audu apstrādes un šūnu kultivēšanas laikā. Mūsu PBS šķīdums ir rūpīgi izstrādāts no augstas tīrības pakāpes sastāvdaļām, lai nodrošinātu stabilitāti un uzticamību katrā eksperimentā. Mūsu PBS šķīduma osmolaritāte un jonu koncentrācija precīzi atbilst cilvēka ķermeņa osmolaritātei un jonu koncentrācijai, tāpēc tas ir izotonisks un nav toksisks vairumam šūnu.
Mūsu PBS šķīduma sastāvs
Mūsu PBS šķīdums ir īpaši tīru fosfātu buferšķīdumu un fizioloģisko šķīdumu maisījums ar pielāgotu pH. 1X darba koncentrācijā tas satur:
8000 mg/l nātrija hlorīda (NaCl)
200 mg/l kālija hlorīda (KCl)
1150 mg/L nātrija fosfāts divbāziskais bezūdens (Na2HPO4)
200 mg/L Kālija fosfāts vienbāziskais bezūdens (KH2PO4)
Šis sastāvs nodrošina optimālu pH un jonu līdzsvaru, kas piemērots plašam bioloģisko lietojumu klāstam.
Mūsu PBS šķīduma lietojumi
Mūsu PBS šķīdums ir ideāli piemērots dažādiem lietojumiem bioloģiskajos pētījumos. Tā izotoniskās un netoksiskās īpašības padara to piemērotu vielu atšķaidīšanai un šūnu konteineru skalošanai. EDTA saturoši PBS šķīdumi ir efektīvi, lai atdalītu pielipušās un saplūdušās šūnas. Tomēr PBS nedrīkst pievienot divvērtīgos metālus, piemēram, cinku, jo tas var izraisīt nogulsnēšanos. Šādos gadījumos ieteicams izmantot Good's buferus. Turklāt mūsu PBS šķīdums ir pieņemama alternatīva vīrusu transportēšanas barotnei RNS vīrusu, tostarp SARS-CoV-2, transportēšanai un uzglabāšanai.
Kvalitātes kontrole
Sterili filtrēts
Uzglabāšana un glabāšanas laiks
Uzglabāt no gaismas pasargātā +2 °C līdz +25 °C temperatūrā.
Pēc atvēršanas uzglabāt 2°C līdz 25°C temperatūrā un izmantot 24 mēnešu laikā.
Pārvadāšanas nosacījumi
Apkārtējā temperatūra
Uzturēšana
Uzglabāt ledusskapī no +2°C līdz +8°C temperatūrā, tumsā. Izvairīties no sasaldēšanas un biežas sasilšanas līdz +37°C, jo tas samazina produkta kvalitāti.
Nesildiet barotni augstāk par 37°C un neizmantojiet nekontrolētus siltuma avotus, piemēram, mikroviļņu krāsnis.
Ja jāizmanto tikai daļa barotnes, pirms lietošanas izņemiet nepieciešamo daudzumu un uzsildiet to līdz istabas temperatūrai.
Sastāvs
Kategorija
Sastāvdaļas
Koncentrācija (mg/l)
Sāls
Kālija hlorīds
200
Kālija fosfāts monobāziskais bezūdens
200
Nātrija hlorīds
8000
Nātrija fosfāts divbāziskais bezūdens
1150

Sākotnēji izstrādāta cilvēka leikēmisko šūnu augšanai suspensijas un monoslāņa kultūrās, RPMI 1640 barotne, pētniekiem un komerciālajiem piegādātājiem veicot modifikācijas, ir kļuvusi piemērota dažādām zīdītāju šūnām. Tā ir īpaši saderīga ar tādām šūnu līnijām kā HeLa, Jurkat, MCF-7, PC12, PBMC, astrocīti un karcinomas.
RPMI 1640 barotne atšķiras no citām šūnu kultūru barotnēm ar savu unikālo sastāvu. Tā satur ievērojamu daudzumu fosfātu, aminoskābju un vitamīnu. Īpaši svarīgi, ka tajā ir biotīns, vitamīns B12 un PABA, kas nav iekļauti Eagle's Minimal Essential Medium vai Dulbecco's Modified Eagle Medium. Turklāt RPMI 1640 barotnē ir ievērojami paaugstināta vitamīnu inozitola un holīna koncentrācija. Tomēr tā nesatur olbaltumvielas, lipīdus vai augšanas faktorus. Tāpēc, lai nodrošinātu optimālus apstākļus šūnu augšanai, parasti ir nepieciešama papildināšana ar 10 % Fetal Bovine Serum (FBS).
RPMI 1640 barotnes buferēšanas sistēma balstās uz nātrija bikarbonātu, un, lai uzturētu fizioloģiski piemērotu pH, tai nepieciešama 5-10 % CO2 vide. Reducējošā aģenta glutationa iekļaušana vēl vairāk atšķir šo barotni no citām.
Kvalitātes kontrole
Sterili filtrēts
Uzglabāšana un derīguma termiņš
Uzglabāt no gaismas pasargātā +2 °C līdz +8 °C temperatūrā.
Pēc atvēršanas uzglabāt 4°C temperatūrā un lietot 6-8 nedēļu laikā.
Pārvadāšanas nosacījumi
Apkārtējā temperatūra
Uzturēšana
Uzglabāt ledusskapī no +2°C līdz +8°C temperatūrā, tumsā. Izvairīties no sasaldēšanas un biežas sasilšanas līdz +37°C, jo tas samazina produkta kvalitāti.
Nesildiet barotni augstāk par 37°C un neizmantojiet nekontrolētus siltuma avotus, piemēram, mikroviļņu krāsnis.
Ja jāizmanto tikai daļa barotnes, pirms lietošanas izņemiet nepieciešamo daudzumu un uzsildiet to līdz istabas temperatūrai.
Sastāvs
Kategorija
Sastāvdaļas
Koncentrācija (mg/l)
Aminoskābes
Glicīns
10.00
L-alanil-L-glutamīns
434.40
L-arginīns
200.00
L-asparagīnsH2O
56.82
L-asparagīnskābe
20.00
L-cistīns 2HCl
65.20
L-glutamīnskābe
20.00
L-Histidīns HClH2O
20.27
L-Hidroksi-L-Prolīns
20.00
L-izoleicīns
50.00
L-leicīns
50.00
L-lizīns HCl
40.00
L-metionīns
15.00
L-fenilalanīns
15.00
L-Prolīns
20.00
L-serīns
30.00
L-treonīns
20.00
L-triptofāns
5.00
L-tirozīns 2Na 2H2O
28.83
L-Valīns
20.00
Vitamīni
p-amīnbenzoskābe
1.00
D-Biotīns
0.20
Holīna hlorīds
3.00
D-kalcija pantotenāts
0.25
Folijskābe
1.00
mio-Inozitols
35.00
Nikotīnamīds
1.00
Piridoksīna HCl
1.00
Riboflavīns
0.20
Tiamīna HCl
1.00
B12 vitamīns
0.005
Neorganiskie sāļi
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Citi komponenti
D-glikoze
2000.00
L-Glutationa reducēts
1.00
Fenola sarkanais nātrija sāls
5.30
, w: 3,1 g/l glikozes, w: 2,5 mM L-glutamīna, w: 15 mM HEPES, w: 0,5 mM nātrija piruvāta, w: 1,2 g/l NaHCO3")
Šī unikālā formula apvieno Dulbecco modificēto Eagle barotni (DMEM) un Ham's F-12 (Ham's barības vielu maisījumu F-12) precīzā 1:1 attiecībā. L-glutamīna pievienošana vēl vairāk uzlabo tās sastāvu.
DMEM, kas ir atvasināts no Eagle's Minimal Essential Medium (EMEM), salīdzinājumā ar savu priekšteci piedāvā paaugstinātu aminoskābju un vitamīnu koncentrāciju. Savukārt Ham's F-12 ir balstīts uz Ham's F-10 barotni, nodrošinot papildu nepieciešamo komponentu kopumu.
Lai atbalstītu optimālu šūnu augšanu, parasti DMEM:Ham's F12 papildina ar FBS tipiskā koncentrācijā 5–10 %. Šī piedeva ir nepieciešama, jo barotnē trūkst augšanas hormonu, lipīdu un proteīnu, kas ir būtiski šūnu attīstībai.
DMEM:Ham's F12 satur pH buferu sistēmu un bieži tiek papildināts ar fenola sarkano, kas ir pH indikators. Kultivētām šūnām DMEM:Ham's F12 vai jebkurā vidē, kurā izmanto bikarbonāta buferu sistēmu, ir nepieciešama kontrolēta CO2 vide 5–10 % līmenī, lai uzturētu atbilstošu pH līmeni.
Kvalitātes kontrole
Sterili filtrēts
Uzglabāšana un derīguma termiņš
Uzglabāt +2 °C līdz +8 °C temperatūrā, aizsargājot no gaismas.
Pēc atvēršanas uzglabāt 4 °C temperatūrā un izlietot 6–8 nedēļu laikā.
Piegādes nosacījumi
Vides temperatūra
Uzglabāšana
Uzglabāt ledusskapī +2 °C līdz +8 °C temperatūrā, aizsargājot no gaismas. Izvairieties no sasaldēšanas un biežas sasilšanas līdz +37 °C, jo tas samazina produkta kvalitāti.
Nesildiet barotni virs 37 °C un nelietojiet nekontrolētus siltuma avotus, piemēram, mikroviļņu krāsnis.
Ja jāizmanto tikai daļa no barotnes, pirms lietošanas izņemiet nepieciešamo daudzumu un uzsildiet to līdz istabas temperatūrai.
Sastāvs
Kategorija
Sastāvdaļas
Koncentrācija (mg/l)
Aminoskābes
Glicīns
18,75
L-alanīns
4,45
L-arginīna HCl
147,50
L-asparagīns H₂O
7,50
L-asparagīnskābe
6,65
L-cisteīns HCl H₂O
17,56
L-cistīns 2 HCl
31,29
L-glutamīnskābe
7,35
L-glutamīns
365,00
L-histidīna HCl H₂O
31,48
L-izoleicīns
54,47
L-leicīns
59,05
L-lizīna HCl
91,25
L-metionīns
17,24
L-fenilalanīns
35,48
L-prolīns
17,25
L-serīns
26,25
L-treonīns
53,45
L-triptofāns
9,02
L-tirozīns 2 Na 2 H2O
55,79
L-valīns
52,85
Vitamīni
D-biotīns
0,0035
Holīna hlorīds
8,98
D-kalcija pantotenāts
2,24
Folijskābe
2,66
mio-inozitols
12,60
Nikotinamīds
2,02
Piridoksīna HCl
0,031
Piridoksāls HCl
2,00
Riboflavīns
0,219
Tiamīna HCl
2,17
Vitamīns B12
0,68
Neorganiskie sāļi
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO3)3 9 H2O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Citi komponenti
D-glikoze
3151,00
Hipoksantīns
2,40
HEPES
3574,50
Linolskābe
0,042
Lipoiskā skābe
0,105
Fenola sarkanais nātrija sāls
8,63
Putrescīns 2 HCl
0,081
Nātrija piruvāts
55,00
Timidīns
0,365
, w: 3,0 g/l glikoze, w: stabils glutamīns, w: 2,0 mM nātrija piruvāts, w: 2,2 g/l NaHCO3")
- 0,02 pie 20-25 °C. Katra partija ir pārbaudīta uz sterilitāti un mikoplazmas un baktēriju neesamību. Uzglabāšana Uzglabāt ledusskapī +2 °C līdz +8 °C temperatūrā, nošķirti no gaismas. Saldēšana un sildīšana līdz +37 °C pazemina produkta kvalitāti. Neuzkarsējiet barotni līdz vairāk nekā 37 °C un neizmantojiet nekontrolējamas siltuma avotus (piemēram, mikroviļņu krāsnis). Ja ir jāizmanto tikai daļa barotnes, izņemiet šo daudzumu no pudeles un sildiet to istabas temperatūrā. Jebkura barotnes, izņemot pamata barotni, glabāšanas termiņš ir 6 līdz 8 nedēļas no atvēršanas brīža. Sastāvs Sastāvdaļas mg/L Neorganiskie sāļiKalcija hlorīds x 2H2O132,00 Magnija sulfāts97,67 Kālija hlorīds400,00 Nātrija hlorīds6460,00 Dinātrija hidrogēnfosfāts (bezūdens)504,00 Citi komponentiD(+)-glikoze (bezūdens)3000 Glutationa (reducēts)0,5 Gaļas peptonis600,00 Fenola sarkanais nātrija sāls11 AminoskābesL-alanīns13,36 L-arginīns x HCl42,14 L-asparagīns x H2O45,03 L-asparagīnskābe19,97 L-cisteīns x HCl x H2O31,75 L-glutamīns (stabils)219,15 L-glutamīnskābe22,07 Glicīns7,51 L-histidīns x HCl x H2O20,96 L-hidroksiprolīns19,67 L-izoleicīns39,36 L-leicīns39,36 L-lizīns x HCl36,54 L-metionīns14,92 L-fenilalanīns16,52 L-prolīns17,27 L-serīns26,28 L-treonīns17,87 L-triptofāns3,06 L-tirozīna dinātrija sāls26,10 L-valīns17,57 Vitamīnip-Aminobenzoīnskābe1,0 Askorbīnskābe0,56 D(+)-biotīns0,20 D-kalcija pantotenāts0,20 Holīna hlorīds5,0 Folijskābe10 mio-inozitols36 Nikotinamīds0,5 Nikotīnskābe0,5 Piridoksāla HCl0,50 Piridoksīna HCl0,50 Riboflavīns0,20 Tiamīna HCl0,20 Vitamīns B122,00

Barotne 199 piedāvā virkni pielietojumu šajā jomā. Ar to var efektīvi uzturēt kumulus-ocītu kompleksu (COC) un atbalstīt olšūnu nobriešanu in vitro. Turklāt to izmanto aspirācijas līniju skalošanai, vācot olšūnas no Vācijas Holšteinas govīm. Turklāt Medium 199 ir lieliska barotne no žurkām iegūtu sirds endotēlija šūnu kultivēšanai. Šie pielietojumi liecina par barotnes 199 daudzpusību un pielāgojamību dažādām eksperimentālām vajadzībām.
Vēsture
Šķīduma Nr. 199 izstrāde pagājušā gadsimta 50. gados iezīmēja ievērojamu progresu audu kultūru barotņu jomā. Pirms tās ieviešanas daudzas barotnes balstījās uz dzīvnieku izcelsmes produktiem un audu ekstraktiem. Tomēr Morgans un kolēģi radīja apvērsumu šajā jomā, formulējot pilnīgi noteiktu barības avotu šūnu kultūrām. Veicot eksperimentus ar dažādām vitamīnu, aminoskābju un citu faktoru kombinācijām, viņi atklāja, ka barotnei Nr. 199 piemīt izcilas augšanu veicinošas īpašības.
Kvalitātes kontrole
pH = 7,2 +/
- 0,02 20-25 °C temperatūrā.
Katra partija ir pārbaudīta attiecībā uz sterilitāti un mikoplazmas un baktēriju neesamību.
Uzturēšana
Uzglabāt ledusskapī +2°C līdz +8°C temperatūrā, tumsā. Sasaldēšana un sasilšana līdz +37 °C samazina produkta kvalitāti.
Nesildiet barotni līdz temperatūrai, kas pārsniedz 37° C, un neizmantojiet nekontrolējamus siltuma avotus (piemēram, mikroviļņu krāsnis).
Ja jāizmanto tikai daļa barotnes, izņemiet šo daudzumu no pudeles un uzsildiet to istabas temperatūrā.
Jebkuras barotnes, izņemot pamata barotni, derīguma termiņš ir 8 nedēļas no izgatavošanas datuma.
Sastāvs
Sastāvdaļas
mg/l
Neorganiskie sāļi
Kalcija hlorīds x 2H2O
264,92
Dzelzs (III) nitrāts x 9H2O
0,72
Magnija sulfāts
97,67
Kālija hlorīds
400,00
Nātrija acetāts x 3H2O
82,95
Nātrija hlorīds
6,800.00
Nātrija dihidrogēnfosfāts x H2O
140,00
Citi komponenti
Adenīna sulfāts
10,00
AMP
0,20
ATP
1,00
Holesterīns
0,20
2'-deoksiriboze
0,50
D(+)-glikoze bezūdens
1,000.00
Glutations (sarkans)
0,05
Guanīns x HCl
0,30
Hipoksantīns
0,30
Fenola sarkanais
10,00
D-riboze
0,50
Timīns
0,30
Tween 80
4,90
Uracils
0,30
Ksantīns
0,30
NaHCO3
2,200.00
Aminoskābes
L-alanīns
25,00
L-arginīns x HCl
70,00
L-asparagīnskābe
30,00
L-cisteīns x HCl x H2O
0,10
L-cistīns
20,00
L-glutamīns stabils
149,00
L-glutamīnskābe
67,00
Glicīns
50,00
L-histidīns x HCl x H2O
21,88
L-hidroksiprolīns
10,00
L-izoleicīns
20,00
L-leicīns
60,00
L-lizīns x HCl
70,00
L-metionīns
15,00
L-fenilalanīns
25,00
L-Prolīns
40,00
L-serīns
25,00
L-treonīns
30,00
L-triptofāns
10,00
L-tirozīns
40,00
L-valīns
25,00
Vitamīni
4-amino benzoskābe
0,05
Askorbīnskābe
0,05
D(+)-biotīns
0,01
Kalciferols
0,10
D-kalcija pantotenāts
0,01
Holīna hlorīds
0,50
Folijskābe
0,01
mio-inositols
0,05
Menadions
0,01
Nikotīnskābe
0.025
Nikotīnamīds
0.025
Piridoksāls x HCl
0.025
Piridoksols x HCl
0.025
Riboflavīns
0,01
DL-α-tokoferola fosfāta dinātrija sāls
0,01
Tiamīns x HCl
0,01
Vitamīna A acetāts
0,14

IMDM ir labi piemērots strauji proliferējošām, augsta blīvuma šūnu kultūrām, tostarp Jurkat, COS-7 un makrofāgu šūnām. Dažādas IMDM modifikācijas, kas pieejamas dažādiem šūnu kultūru lietojumiem, var atrast, izmantojot barotņu atlases rīku. Šķidrās barotnes nodrošina būtiskas barības vielas visiem šūnu kultūru lietojumiem. Katra no mūsu augstas kvalitātes šūnu kultūru barotnēm tiek ražota saskaņā ar sākotnēji publicēto formulu vai modifikācijām, kas nepieciešamas, lai nodrošinātu kultūras barotnes nemainīgu veiktspēju un stabilitāti.
IMDM pret DMEM
IMDM dzelzs nitrāta vietā ir kālija nitrāts, kā arī HEPES un nātrija piruvāts. IMDM papildu komponenti padara to piemērotāku specializētiem šūnu tipiem un specifiskiem lietojumiem nekā DMEM.
IMDM pret RPMI
IMDM un RPMI ir atšķirīgi sastāvi, kas var būt svarīgi PMA/jonomicīna stimulācijai. Viena būtiska atšķirība ir Ca2+ koncentrācija. RPMI satur 0,42 mM Ca2+, bet IMDM
- 1,49 mM.
Kvalitātes kontrole
pH = 7,2 +/
- 0,02 20-25 °C temperatūrā.
Katra partija ir pārbaudīta attiecībā uz sterilitāti un mikoplazmas un baktēriju neesamību.
Uzturēšana
Uzglabāt ledusskapī +2°C līdz +8°C temperatūrā tumsā. Sasaldēšana un sasilšana līdz +37 °C samazina produkta kvalitāti.
Nesildiet barotni līdz temperatūrai, kas pārsniedz 37° C, un neizmantojiet nekontrolējamus siltuma avotus (piemēram, mikroviļņu krāsnis).
Ja jāizmanto tikai daļa barotnes, izņemiet šo daudzumu no pudeles un uzsildiet to istabas temperatūrā.
Jebkuras barotnes, izņemot pamata barotni, derīguma termiņš ir 8 nedēļas no izgatavošanas datuma.
Sastāvs
Sastāvdaļas
mg/l
Neorganiskie sāļi
Kalcija hlorīds x 2 H2O
219,00
Kālija hlorīds
330,00
Kālija nitrāts
0.076
Bezūdens magnija sulfāts
97,73
Nātrija hlorīds
4,505.00
Bezūdens nātrija dihidrogēnfosfāts
109,00
Nātrija selenīts
0,02
Citi komponenti
D(+)-glikoze bezūdens
4,500.00
HEPES
5,958.00
Nātrija piruvāts
110,00
Fenola sarkanais
15,00
Aminoskābes
L-alanīns
25,00
L-arginīns x HCl
84,00
L-asparagīns x H2O
25,00
L-asparagīnskābe
30,00
L-cistīns x 2HCl
91,24
L-glutamīns
584,00
L-glutamīnskābe
75,00
Glicīns
30,00
L-histidīns x HCl x H2O
42,00
L-izoleicīns
104,80
L-leicīns
104,80
L-lizīns x HCl
146,20
L-metionīns
30,00
L-fenilalanīns
66,00
L-Prolīns
40,00
L-serīns
42,00
L-treonīns
95,20
L-triptofāns
16,00
L-tirozīns x 2Na
104,20
L-valīns
93,60
Vitamīni
D(+)-biotīns
0.013
D-kalcija pantotenāts
4,00
Holīna hlorīds
4,00
Folijskābe
4,00
mio-inositols
7,20
Šūnu kultūras barotnes: pārskats
Dabaszinātņu jomā viena no svarīgākajām metodēm ir šūnu kultivēšana. Ar terminu „šūnu kultivēšana” saprot šūnu, audu vai orgānu izņemšanu no dzīvnieka vai auga un to turpmāku implantēšanu mākslīgā vidē, kas ir labvēlīga to izdzīvošanai un/vai augšanai. Optimālai šūnu attīstībai nepieciešamās pamatvides prasības ir kontrolēta temperatūra, substrāts šūnu piestiprināšanai, atbilstoša augšanas vide un inkubators, kas uztur optimālu pH līmeni un osmolalitāti. Lai šūnas varētu pilnībā attīstīties, tām ir jānodrošina šie apstākļi.
Atbilstošas augšanas vides izvēle in vitro kultivēšanai ir šis šūnu kultūras posms, kas ir gan vissvarīgākais, gan izšķirošais. Augšanas vide, ko dēvē arī par kultūras vidi, ir šķidrums vai gēls, kas izstrādāts, lai veicinātu organismu attīstību mikroskopiskā, šūnu vai augu līmenī. Šūnu kultivēšanai izmantotā vide bieži satur pietiekamu enerģijas daudzumu un vielas, kas regulē šūnu ciklu. Kultūras vides galvenās sastāvdaļas ir aminoskābes, vitamīni, neorganiskie sāļi, glikozes un serums. Serums tiek pievienots videi, jo tas darbojas kā augšanas faktoru, hormonu un saistīšanās faktoru avots. Papildus barības vielu nodrošināšanai vide palīdz arī uzturēt pH un osmolalitātes līmeni.
Šūnu kultūrā izmantoto barotņu veidi
Gan cilvēka, gan dzīvnieku šūnas var audzēt gan mākslīgā vai sintētiskā barotnē, gan pilnīgi dabīgā barotnē, kas papildināta ar dabīgiem elementiem. Turpinājumā sniegsim pārskatu par dažādiem pašlaik pieejamiem barotņu veidiem.
Dabiskās barotnes
Dabiskajā barotnē var atrast tikai bioloģiskos šķidrumus, kas pastāv to dabiskajā stāvoklī. Dabiskās barotnes ir ļoti noderīgas un viegli izmantojamas dažādu dzīvnieku šūnu tipu kultivēšanai. Nepietiekama izpratne par precīziem komponentiem, kas veido dabisko barotni, ir galvenais faktors, kas ietekmē zemo rezultātu atkārtojamību, izmantojot dabiskās barotnes.
Mākslīgās barotnes
Mākslīgo vai sintētisko barotņu sagatavošana ietver barības vielu (gan organisko, gan neorganisko), seruma proteīnu, ogļhidrātu, kofaktoru, vitamīnu un sāļu pievienošanu, kā arī O2 un CO2 gāzes fāžu pievienošanu [1].
Ir izstrādāti dažādi mākslīgo barotņu veidi, lai pildītu vienu vai vairākas no šādām funkcijām: 1) Tūlītēja izdzīvošana (līdzsvarots sāļu šķīdums ar precīzu pH un osmotisko spiedienu). 2) Ilgstoša izdzīvošana (līdzsvarots sāļu šķīdums, kam pievienotas dažādas organisko ķīmisko vielu un/vai seruma formulācijas). 3) Neierobežota attīstība. 4) Specializētas funkcijas.
Mākslīgajiem barotnēm ir četras atšķirīgas klasifikācijas:
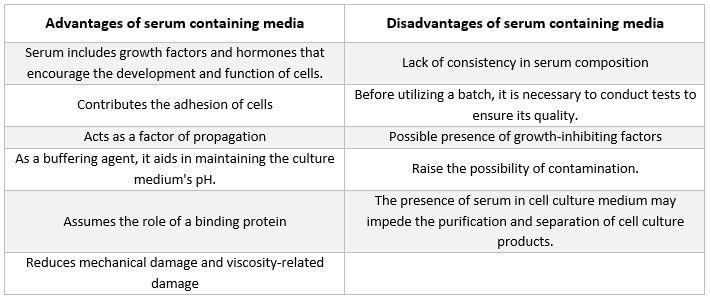
Serumu saturošas barotnes
Visbiežāk sastopamais piedevas veids dzīvnieku šūnu audzēšanai izmantotajā barotnē ir liellopu augļa serums. To pievieno kultūras barotnei kā lētu piedevu, lai panāktu iespējami labākos augšanas apstākļus. Papildus tam, ka serums darbojas kā nestabilu vai ūdenī nešķīstošu uzturvielu, hormonu un augšanas faktoru, proteāžu inhibitoru un citu vielu pārvadātājs vai helators, tas arī saista un neitralizē kaitīgas molekulas.
Seruma nesaturošas barotnes
Seruma klātbūtnei barotnēs ir vairāki trūkumi, un tā var izraisīt būtiskas interpretācijas kļūdas imunoloģiskajos pētījumos [2, 3]. Ir izstrādātas dažādas bezseruma barotnes [4, 5]. Šīs barotnes parasti ir speciāli izstrādātas, lai atbalstītu viena šūnu tipa kultivēšanu, piemēram, „Knockout Serum Replacement” un „Knockout DMEM” no „Thermo Fisher Scientific”, kā arī „mTESR” barotne no „Stem Cell Technologies” [6], kas paredzēta cilmes šūnām [7].
Turklāt šajās barotnēs ir iekļauti noteikti daudzumi attīrītu augšanas faktoru, lipoproteīnu un citu proteīnu, kurus citādi parasti nodrošina serums [8]. Šīs barotnes bieži dēvē par „definētām kultivēšanas barotnēm”, jo to sastāvdaļas ir labi izpētītas.
Ķīmiski definētas barotnes
Šīs barotnes satur ultra-tīras neorganiskas un organiskas sastāvdaļas, kas nav piesārņotas ar nekāda veida piemaisījumiem. Tās var saturēt arī tīras olbaltumvielu piedevas, piemēram, augšanas faktorus.
Baktēriju vai rauga ģenētiskā modifikācija kopā ar konkrētu taukskābju, vitamīnu, holesterīna un aminoskābju pievienošanu nodrošina to sastāvdaļu ražošanu [9].
Proteīnu nesaturošas barotnes
Proteīnu nesaturošas barotnes ir tādas, kurās vispār nav proteīnu, bet ir tikai neproteīna elementi. Salīdzinot ar barotnēm, kurām pievienots serums, barotņu izmantošana bez proteīnu piedevas veicina lielāku šūnu proliferāciju un proteīnu ekspresiju, kā arī atvieglo jebkura turpmākajā procesā iegūtā produkta attīrīšanu [10–12]. Proteīns nav iekļauts tādos barotņu sastāvos kā MEM un RPMI-1640. Tomēr nepieciešamības gadījumā var pievienot proteīna piedevu.
Kultivēšanas barotnes un to pamatkomponenti
Komerciālo kultivēšanas barotni var iegādāties pulvera vai šķidrā veidā, un tā bieži satur dažādus uzturvielu avotus, piemēram, aminoskābes, glikozes, sāļus, vitamīnus un citus uztura bagātinātājus.
Šo sastāvdaļu nepieciešamība katrai šūnu līnijai ir atšķirīga, un šīs atšķirības nosaka plašo barotņu sastāvu dažādību. Katrai sastāvdaļai ir sava funkcija, kas tiks izklāstīta turpmākajos punktos:
Buferizācijas sistēmas
Lai uzturētu optimālus augšanas apstākļus, ir jākontrolē pH līmenis, ko bieži vien veic, izmantojot vienu no divām buferizācijas sistēmām:
Dabiskā buferizācijas sistēma
CO₂/H₂CO₃ attiecība atmosfērā ir vienāda ar to barotnē, tādējādi radot dabisku buferizācijas mehānismu. Lai saglabātu šo dabisko buferizācijas mehānismu, kultūras jātur gaisa vidē ar 5–10 % CO₂ saturu, ko bieži panāk, izmantojot CO₂ inkubatoru. Viena no lielākajām dabiskā bufera priekšrocībām ir tā lētums un drošība.
HEPES
Ķīmiskā buferēšana, izmantojot zwitterjonu HEPES, nodrošina lielāku buferēšanas spēju pH diapazonā no 7,2 līdz 7,4 un neprasa regulētu gāzes vidi. Konkrētiem šūnu tipiem lielāka HEPES deva var būt kaitīga. HEPES saturošas barotnes ir arī daudz jutīgākas pret fluorescējošās gaismas fototoksisko iedarbību [13].
Fenolsarkans
pH indikators fenola sarkans bieži tiek iekļauts komerciāli pieejamās kultivēšanas vidēs, ļaujot nepārtraukti uzraudzīt pH. Šīm šūnām vairojoties, to ražotie metabolīti izraisa pH izmaiņas un tādējādi arī vides krāsas maiņu. Fenola sarkanajam ir divkārša ietekme uz barotnes krāsu — skābā pH vidē tā kļūst dzeltena, bet sārmainā pH vidē — violeta. pH 7,4, kas ir optimālā vērtība šūnu kultūrām, liek barotnei izskatīties fluorescējoši sarkanai.
Tomēr fenola sarkanajam ir arī daži trūkumi: pirmkārt, fenola sarkanais spēj imitēt vairāku steroīdu hormonu, galvenokārt estrogēna, darbību [14]. Tāpēc, pētot pret estrogēnu jutīgas šūnas, piemēram, krūšu audus, ieteicams izmantot barotni bez fenola sarkanā. Vairākos bezseruma preparātos fenola sarkana klātbūtne traucē nātrija un kālija līdzsvaru. Šo efektu var neitralizēt, barotnei pievienojot serumu vai liellopu hipofīzes hormonu [15]. Treškārt, fenola sarkana klātbūtne apgrūtina noteikšanu plūsmas citometrijas eksperimentos.
Neorganiskie sāļi
Barotnes, kas satur neorganiskos sāļus, piemēram, nātrija, kālija un kalcija jonus, palīdz uzturēt osmotisko līdzsvaru un regulēt membrānas potenciālu.
Aminoskābes
Tā kā aminoskābes ir proteīnu pamatsastāvdaļas, tās ir būtiska sastāvdaļa ikvienā šūnu audzēšanas barotnē, kas jebkad ir izstrādāta. Tā kā šūnas nespēj pašas ražot noteiktas aminoskābes, ir svarīgi, lai kultūras barotnē būtu iekļautas neaizvietojamās aminoskābes. Tās ir nepieciešamas šūnu vairošanās procesam, un to koncentrācija nosaka maksimālo šūnu blīvumu, ko var sasniegt. Īpaši svarīga ir neaizvietojamā aminoskābe L-glutamīns.
L-glutamīns darbojas kā sekundārs enerģijas avots vielmaiņai un piegādā slāpekli NAD, NADPH un nukleotīdu sintēzei. Tā kā L-glutamīns ir nestabila aminoskābe, kas laika gaitā pārveidojas formā, kuru šūnas nespēj izmantot, tā ir jāpievieno barotnei.
Turklāt barotnē var pievienot neesamās aminoskābes, lai atjaunotu tās, kas iztērētas augšanas procesa laikā. Šūnu augšana tiek veicināta un to dzīvotspēja palielinās, ja augšanas barotnei pievieno neesamās aminoskābes.
Ogļhidrāti
Ogļhidrāti cukuru veidā ir galvenais enerģijas avots. Daudzos barotnēs papildus izplatītākajiem cukuriem — glikozēm un galaktozēm — ir iekļauta arī maltoze un fruktoze.
Olbaltumvielas un peptīdi
Albumīns, transferīns un fibronektīns ir visbiežāk izmantotie proteīni un peptīdi. Tie ir īpaši nozīmīgi barotnēs, kurās nav iekļauts serums. Albumīns, transferīns, aprotinīns, fetuīns un fibronektīns ir daži no proteīniem, kas var atrasties serumā, kas ir bagāts proteīnu avots.
Albumīns ir galvenais asinīs sastopamais proteīns, un tā funkcija ir saistīt un transportēt dažādas vielas, tostarp ūdeni, sāļus, brīvās taukskābes, hormonus un vitamīnus, starp dažādiem orgāniem un šūnām. Albumīna spēja saistīties ar ķīmiskajām vielām padara to par efektīvu līdzekli kaitīgo savienojumu noņemšanai no barotnes, kurā tiek kultivētas šūnas.
Aprotinīns ir aizsarglīdzeklis šūnu kultivēšanas sistēmās, jo tas ir stabils neitrālā un skābā pH vidē, kā arī izturīgs pret augstām temperatūrām un proteolītisko enzīmu izraisītu noārdīšanos. Tas spēj inhibēt virkni serīna proteāžu, tostarp tripsīnu un citas.
Fetuīns ir glikoproteīns, ko dzīvnieku augļu un jaundzimušo dzīvnieku serumā var konstatēt lielākos daudzumos nekā pieaugušo dzīvnieku serumā. Turklāt tas darbojas kā serīna proteāzes inhibitors. Proteīns fibronektīns ir būtiska sastāvdaļa šūnu adhezijas procesā. Transferīns ir olbaltumviela, kas transportē dzelzi un ir atbildīga par dzelzs piegādi šūnu membrānām.
Taukskābes un lipīdi
Tām ir izšķiroša nozīme bezseruma vidē, ja serums nav klāt.
Vitamīni
Šūnu attīstībai un vairošanās procesam ir nepieciešami daudzi vitamīni. Šūnas nespēj ražot pietiekamu vitamīnu daudzumu, tāpēc audu kultūrās tie ir nepieciešami kā uztura bagātinātāji.
Šūnu kultūrā serums ir galvenais vitamīnu avots; tomēr barotnes tiek papildinātas arī ar dažādiem vitamīniem, lai tās būtu piemērotas konkrētam šūnu tipam. Visbiežāk augšanas stimulēšanai izmanto B grupas vitamīnus.
Mikroelementi
Ķīmiskie elementi, piemēram, varš, cinks, selēns un trikarboksilskābes starpprodukti, ir pazīstami kā mikroelementi. Mikroelementus bieži pievieno barotnēm, kurās nav seruma, lai aizstātu tos, kas parasti atrodas serumā. Šie elementi ir svarīgas ķīmiskas sastāvdaļas, kas nepieciešamas veselīgai šūnu attīstībai. Daudzas bioķīmiskās reakcijas, piemēram, fermentu aktivitāte, ir atkarīgas no noteiktiem mikroelementiem.
Barotnes piedevas
Pilnvērtīgajai augšanas barotnei, kas ieteikta noteiktām šūnu līnijām, nepieciešamas papildu sastāvdaļas, kuras nav iekļautas pamata barotnē un serumā. Šie uztura bagātinātāji veicina šūnu augšanu un atbilstošu vielmaiņas funkciju.
Lai gan hormoni, augšanas faktori un signālmolekulas ir būtiski konkrētu šūnu līniju pareizai proliferācijai, vienmēr jāievēro šādi piesardzības pasākumi: tā kā piedevu pievienošana var mainīt pilnās augšanas vides osmolalitāti, kas var kavēt šūnu attīstību, vienmēr ir ieteicams pārbaudīt osmolalitāti pēc piedevu pievienošanas. Lielākajai daļai šūnu līniju optimālā osmolalitāte ir no 260 līdz 320 mOSM/kg.
Antibiotikas
Antibiotikas bieži izmanto, lai kavētu baktēriju un sēnīšu piesārņotāju attīstību [16], lai gan tās nav nepieciešamas šūnu augšanai. Tā kā antibiotikas var slēpt mikoplazmu un rezistentu baktēriju izraisītu piesārņojumu, to rutīna lietošana šūnu kultūrās nav ieteicama [17, 18].
Turklāt antibiotikas var traucēt paaugstinātas jutības šūnu vielmaiņu. Bieži tiek izmantotas MilliporeSigma un Life Technologies ražotās penicilīna–streptomicīna kombinācijas. Plasmocin ir izmantots gliomas šūnu līniju TS603, TS516 un BT260 kultivēšanā [19], un ir pierādīts, ka tas efektīvi novērš mikoplazmas piesārņojumu (20).
Serums
Serumā ir sastopami albumīni, augšanas faktori un augšanas inhibitori. Serums ir viena no vissvarīgākajām šūnu kultūras barotnes sastāvdaļām, jo tas nodrošina aminoskābes, olbaltumvielas, vitamīnus (īpaši taukos šķīstošos vitamīnus, piemēram, A, D, E un K), ogļhidrātus, lipīdus, hormonus, augšanas faktorus, minerālvielas un mikroelementus.
Lai veicinātu kultivēto šūnu attīstību, bieži izmanto serumu, kas iegūts no liellopu augļiem un teļiem. Augļu serums ir bagāts augšanas faktoru avots un ir piemērots šūnu klonēšanai un jutīgu šūnu attīstībai. Tā kā teļu seruma spēja veicināt augšanu ir samazināta, to izmanto kontakta inhibīcijas eksperimentos. Standarta augšanas barotnēs bieži vien ir iekļauts 2–10 % seruma. Seruma pievienošana kultivēšanas barotnei kalpo šādiem mērķiem [21]:
-
Serums nodrošina šūnām nepieciešamos uzturvielu avotus (gan šķīdumā, gan saistīti ar olbaltumvielām).
-
Serumā ir vairāki augšanas faktori un hormoni, kas piedalās augšanas veicināšanā un specializētajā šūnu darbībā.
-
Tajā ir daudz saistošo proteīnu, piemēram, albumīns un transferīns, kas transportē citas ķīmiskās vielas uz šūnu. Piemēram, albumīns piegādā taukus, vitamīnus, hormonus utt. uz šūnām.
-
Tas nodrošina arī proteīnus, piemēram, fibronektīnu, kas palielina šūnu saistīšanos ar substrātu. Turklāt tas rada izplatīšanās elementus, kas palīdz šūnu izplešanās procesā pirms dalīšanās.
-
Tas piegādā proteāžu inhibitorus, kas novērš proteolīzi šūnās.
-
Tas satur arī minerālvielas, piemēram, Na+, K+, Zn2+ un Fe2+.
-
Tas palielina barotnes viskozitāti, tādējādi aizsargājot šūnas no mehāniskiem bojājumiem suspensijas kultūras maisīšanas laikā.
-
Tas darbojas arī kā buferis.
Atsauces
[1] Morgan J, Morton H, Parker R. Dzīvnieku šūnu barošana audu kultūrā; sākotnējie pētījumi par sintētisko barotni. Proc Soc Exp Biol Med. 1950;73:1-8
[2] Kerbel R, Blakeslee D. Teļa augļa seruma komponenta ātra adsorbcija kultivētajās zīdītāju šūnās. Potenciāls artefaktu avots pētījumos par antiserumiem pret šūnu specifiskiem antigēniem. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. Seruma pievienošana barotnei, ko izmanto šūnu suspensiju sagatavošanai, kā iespējamais artefaktu avots šūnu mediētajās reakcijās, kuras pētītas, izmantojot popliteālā limfmezgla testu. J Immunogenet. 1980;7:483-9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Komerciālas bezseruma barotnes: hibridomu augšana un monoklonālo antivielu ražošana. J Immunol Methods. 1991;145:175–83
[5] Barnes D, Sato G. Metodes kultivēto šūnu audzēšanai bezseruma barotnē. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O, u.c. HSP70 šaperoni nogādā RNS-brīvo TDP-43 anizotropiskās intranukleārās šķidrās sfēriskās apvalkos. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, u.c. Dauna sindroma izraisītā novecošanās traucē neironu priekšteču kodola arhitektūru. Cell Stem Cell. 2022;29:116–130.e7
[8] Iscove N, Melchers F. Seruma pilnīga aizstāšana ar albumīnu, transferīnu un sojas lipīdiem lipopolisaharīdiem reaģējošu B limfocītu kultūrās. J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Hibridomu augšanai un monoklonālo antivielu ražošanai paredzētas ķīmiski definētas bezproteīnu barotnes sistemātiska uzlabošana. J Biotechnol. 1996;45:111-23
[10] Darfler F. Proteīnu nesaturoša barotne hibridomu un citu imūnsistēmas šūnu audzēšanai. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Šeruma nesaturoša šūnu kultūra: vienota pieeja. Cell. 1980;22:649–55
[12] Hamilton W, Ham R. Ķīniešu kāmju šūnu līniju klonālā audzēšana proteīnu nesaturošās barotnēs. In Vitro. 1977;13:537–47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Gaismas iedarbībai pakļautas HEPES saturošas kultūras vides citotoksiskās iedarbības analīze. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. Fenolsarkanais audu kultūras barotnēs ir vājš estrogēns: sekas estrogēniem reaģējošu šūnu pētījumiem kultūrā. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Bezseruma barotņu izstrāde. In: Master JRW, redaktors. Dzīvnieku šūnu kultūras, 3. izdevums. Oksforda: Oxford University Press; 2000.
[16] Perlman D. Antibiotiku lietošana šūnu kultūras barotnēs. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. Mikoplazmu infekcijas izplatība un kontrole šūnu kultūrās. In Vitro. 1976;12:643–8
[18] Masters J, Stacey G. Barotnes nomaiņa un šūnu līniju pārstādīšana. Nat Protoc. 2007;2:2276–84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M, et al. Histonu demetilāze KDM6A tieši uztver skābekli, lai regulētu hromatīnu un šūnu likteni. Science. 2019;363:1217–1222
[20] Molla Kazemiha V., Azari S., Amanzadeh A., Bonakdar S., Shojaei Moghadam M., Habibi Anbouhi M, u.c. Plasmocin™ efektivitāte dažādās ar mollicutes inficētās zīdītāju šūnu līnijās salīdzinājumā ar šūnu kultūrās parasti lietotajām antibiotikām: vietējā pieredze. Cytotechnology. 2011;63:609-20
[21] Kragh Hansen U. Liganda saistīšanās ar seruma albumīnu molekulārie aspekti. Pharmacol Rev. 1981;33:17-53