
, L-alanyl-L-glutaminløsning")
Volum: 100 ml
Oppbevaring: ≤ –15 °C
Sterilitet: Sterilfiltrert
Stabil glutaminløsning (L-alanyl-L-glutamin, 200 mM) er en svært stabil dipeptidform av L-glutamin som er utviklet som en direkte erstatning for konvensjonell L-glutamin i cellekulturmedier. L-glutamin er en essensiell aminosyre og en viktig energikilde for dyrkede celler, og spiller en avgjørende rolle i cellevekst, metabolisme og proteinsyntese.
Anvendelse og fordeler
I standard flytende medier brytes L-glutamin relativt raskt ned ved 37 °C, noe som fører til dannelse av giftige biprodukter som ammoniumioner, som kan påvirke cellenes levedyktighet og eksperimentelle resultater negativt. Stabil glutamin overvinner denne begrensningen ved å tilby en ikke-nedbrytbar dipeptidform som forblir intakt under dyrkningsforhold.
Cellene spalter dipeptidbindingen enzymatisk for å frigjøre L-glutamin etter behov, noe som sikrer en kontinuerlig tilførsel av fersk glutamin samtidig som det forhindrer opphopning av skadelige avfallsprodukter. Dette gjør løsningen spesielt fordelaktig for langvarige cellekulturer og vekstsystemer med høy tetthet.
Formulering og bruk
L-Alanyl-L-Glutamin-løsningen tilberedes i vann av cellekulturkvalitet i en konsentrasjon på 200 mM og er sterilt filtrert for kontamineringsfølsomme anvendelser. Den kan fortynnes direkte i komplette medier i henhold til eksperimentelle krav. Oppbevares ved ≤ –15 °C og unngå gjentatte fryse-tine-sykluser for å opprettholde produktets stabilitet.
Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
")
Volum: 100 ml
Oppbevaring: +2 °C til +8 °C
Sterilitet: Sterilfiltrert
HEPES-bufferløsning (1 M), også kjent som N-2-hydroksyetylpiperazin-N-2-etansulfonsyre, er et zwitterionisk organisk buffermiddel som er mye brukt i cellekulturmedier. Det er utviklet for å opprettholde stabile pH-forhold i det fysiologiske området 6,7 til 8,6, og støtter optimal cellefunksjon under in vitro-applikasjoner.
Anvendelse og fordeler
HEPES gir pålitelig bufferkapasitet i cellekultursystemer, spesielt når celler håndteres utenfor en CO₂-inkubator. Tilsetning av 10–25 mM HEPES til dyrkningsmedier gir forbedret pH-stabilitet under lengre manipulasjonsperioder, noe som bidrar til å opprettholde konsistente eksperimentelle forhold.
Denne bufferen er membranimpermeabel, har minimal innvirkning på biokjemiske reaksjoner og viser sterk kjemisk og enzymatisk stabilitet. Disse egenskapene gjør den egnet for et bredt spekter av cellekultur
- og biokjemiske anvendelser.
Formulering og bruk
Løsningen leveres som et 1 M konsentrat tilberedt i vann av cellekulturkvalitet og er sterilt filtrert for bruk i kontamineringsfølsomme miljøer. Den kan fortynnes til ønsket arbeidskonsentrasjon avhengig av bruksområdet. Oppbevares ved +2 °C til +8 °C og håndteres under aseptiske forhold for å bevare produktets integritet.
Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
, 250 g/l")
Volum: 50 ml
Oppbevaring: +2 °C til +8 °C
Sterilitet: Sterilfiltrert
D-(+)-glukoseoppløsning (dekstroseoppløsning) er et sterilt, bruksklart kosttilskudd som inneholder det naturlig forekommende sukkeret D-(+)-glukose, en sentral komponent i cellemetabolismen. Glukose er involvert i viktige biologiske prosesser som energiproduksjon, glykosylering og dannelse av glykaner som bidrar til cellestruktur og -funksjon.
Anvendelse og fordeler
Denne glukoseoppløsningen er mye brukt som tilskudd i cellekulturmedier og i en rekke celle
- og molekylærbiologiske anvendelser. Som primær karbon
- og energikilde støtter glukose cellevekst, proliferasjon og metabolsk aktivitet. Dens involvering i biosyntetiske veier gjør den også avgjørende for å opprettholde normal cellefysiologi og eksperimentell konsistens.
Formulering og bruk
Løsningen leveres i en høy konsentrasjon på 250 g/l glukose, noe som gjør det mulig å fortynne den fleksibelt i dyrkningsmedier i henhold til eksperimentelle krav. Den er sterilt filtrert for å sikre at den er egnet for kontamineringsfølsomme anvendelser. Oppbevares ved +2 °C til +8 °C og håndteres aseptisk for å opprettholde produktkvaliteten og stabiliteten.
Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
, oppløsning 100x")
Volum: 10 ml
Oppbevaring: +2 °C til +8 °C
Sterilitet: Sterilfiltrert
Insulin-Transferrin-Selenium (ITS) Solution (100x) er et kjemisk definert supplement utviklet for et bredt spekter av cellekulturapplikasjoner. Det brukes oftest som et tilsetningsstoff til basale cellekulturmedier for å støtte cellevekst under reduserte serum
- eller serumfrie forhold.
Anvendelse og fordeler
Vårt ITS-tilskudd gir essensielle komponenter som kreves for optimal ytelse av serumfrie medier. Ved å supplere konvensjonelle næringsmedier med ITS kan behovet for føtalt bovint serum (FBS) for rutinemessig vedlikehold av mange cellelinjer reduseres betydelig. Dette bidrar til å minimere variasjon knyttet til bruk av serum, samtidig som man opprettholder konsistent cellevekst og levedyktighet.
Insulin støtter cellulær opptak og metabolisme av viktige næringsstoffer, transferrin letter jerntransport, og selen bidrar til antioksidantforsvar og enzymatisk aktivitet. Sammen fremmer disse komponentene balansert cellemetabolisme og forbedret reproduserbarhet i definerte kultursystemer.
Sammensetning og bruk
Insulin-Transferrin-Selenium (ITS) leveres som en 100 ganger konsentrert løsning i Earle's Balanced Salt Solution (EBSS) uten fenolrødt. For standardanvendelser fortynnes det 1:100 i det aktuelle basismediet for å oppnå den anbefalte arbeidskonsentrasjonen. Oppbevares ved +2 °C til +8 °C og håndteres under aseptiske forhold for å opprettholde produktets stabilitet og sterilitet.
Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
")
Volum: 5 ml
Oppbevaring: +2 °C til +8 °C
Sterilitet: Sterilfiltrert
Insulin Human Recombinant Solution er et kjemisk definert tilskudd som ofte brukes til dyrking av pattedyrcellelinjer, inkludert Chinese Hamster Ovary (CHO)-celler. Denne løsningen av cellekulturkvalitet inneholder rekombinant humant insulin uttrykt i Saccharomyces cerevisiae, noe som sikrer høy renhet og jevn ytelse i forskningsapplikasjoner.
Anvendelse og fordeler
Insulin tilsettes rutinemessig i serumfrie og kjemisk definerte medier for å fremme cellevekst og produktivitet. Som et viktig regulerende hormon støtter insulin cellulær opptak, utnyttelse og lagring av glukose, aminosyrer og fettsyrer. Det hemmer også nedbrytningen av glykogen, proteiner og lipider, og bidrar dermed til forbedret cellelevbarhet og metabolsk stabilitet i dyrkningssystemer. Den kjemisk definerte formuleringen støtter reproduserbarhet og minimerer variasjon i sensitive cellekulturarbeidsflyter.
Biologiske egenskaper og bruk
Insulin er et to-kjede polypeptidhormon som produseres naturlig av β-cellene i bukspyttkjertelens øyer. Det har en molekylvekt på omtrent 5 800 Da. α
- og β-kjedene er koblet sammen av to disulfidbindinger mellom kjedene, og α-kjeden inneholder én disulfidbinding innenfor kjeden. For cellekulturformål bør løsningen håndteres under aseptiske forhold og oppbevares ved +2 °C til +8 °C for å opprettholde stabilitet og ytelse.
Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
")
Volum: 100 ml
Oppbevaring: +2 °C til +8 °C
Sterilitet: Sterilfiltrert
Natriumpyruvatløsning (100 mM) er et sterilt, bruksklart tilskudd som er utviklet for å gi en ekstra, lett tilgjengelig energikilde for cellekulturmedier. Natriumpyruvat spiller en viktig rolle i cellenes energimetabolisme og støtter veksten av metabolsk aktive og raskt prolifererende celler, for eksempel tumorceller. Tilskudd kan forbedre cellenes levedyktighet og bidra til å opprettholde metabolsk stabilitet i kultursystemer.
Anvendelse og fordeler
Denne løsningen er mye brukt i rutinemessig cellekultur for å berike medier med pyruvat og fremme optimale vekstforhold. Den støtter ATP-produksjon, kan bidra til å redusere oksidativt stress og bidrar til forbedret metabolsk ytelse hos dyrkede celler. Produktet er produsert i vann av cellekulturkvalitet og sterilt filtrert, og sikrer jevn kvalitet og reproduserbarhet i forskningsarbeidsflyter.
Bruk og kompatibilitet
Den anbefalte sluttkonsentrasjonen for de fleste cellekulturformål er 1 mM natriumpyruvat, oppnådd ved å fortynne 100 mM stamløsningen 1:100 i komplett dyrkningsmedium. Løsningen er kompatibel med et bredt spekter av basale medier og pattedyrcellelinjer. Oppbevares ved +2 °C til +8 °C og beskyttes mot forurensning for å opprettholde produktets stabilitet.
Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
")
Volum: 100 ml Oppbevaring: ≤-15 °C Sterilitet: Sterilfiltrert
Antibiotic/Antimycotic Solution (100x) er et sterilt, bruksklart konsentrat som er utviklet for å redusere risikoen for mikrobiell kontaminering i cellekulturer og relaterte laboratorieapplikasjoner. Denne 100x-løsningen inneholder en veletablert kombinasjon av penicillin, streptomycin og amfotericin B, som gir bredspektret antimikrobiell aktivitet mot grampositive og gramnegative bakterier, gjærsopp og filamentøse sopper. Formuleringen er egnet for bruk i eukaryote cellekulturer, bakteriemedier og andre systemer som er følsomme for kontaminering, og støtter rene og konsekvente laboratorieoperasjoner.
Anvendelse og fordeler Denne løsningen er optimalisert for rutinemessige forskningsprotokoller og brukes i stor utstrekning for å opprettholde aseptiske forhold i arbeidsflyter for cellekulturer. Den gir pålitelig ytelse i kontamineringssensitive miljøer, og hjelper forskere med å redusere risikoen for mikrobiell overvekst uten at det går på bekostning av cellenes helse eller eksperimentell reproduserbarhet. Den sterilfiltrerte formuleringen eliminerer behovet for ytterligere solubiliseringstrinn, noe som bidrar til strømlinjeformet tilberedning av medier og reduserer variabiliteten i daglige laboratorieprosedyrer.
Bruk og kompatibilitet For å oppnå standard arbeidskonsentrasjoner fortynner du løsningen 1:100 i ditt komplette dyrkingsmedium. Produktet er kompatibelt med et bredt spekter av pattedyrcellelinjer og basalmedier. Med konsekvent lagertilgjengelighet kan forskere dra nytte av pålitelig forsyningskontinuitet og forenklet logistikkplanlegging. Løsningen skal oppbevares ved ≤ -15 °C og beskyttes mot gjentatte fryse-tine-sykluser for å opprettholde stabiliteten. Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.
")
Volum: 100 ml Oppbevaring: +2 °C til +8 °C Sterilitet: Sterilfiltrert
MEM Non-Essential Amino Acids (100x) er et sterilt tilskudd som er utviklet for å forbedre cellevekst og levedyktighet i cellekultursystemer fra pattedyr. Formuleringen tilsvarer et 100x konsentrat av de ikke-essensielle aminosyrene som finnes i standard Minimum Essential Medium (MEM), noe som muliggjør direkte tilskudd til basalmedier med minimal forberedelse.
Anvendelse og fordeler Dette tilskuddet gir en ekstra aminosyrepool for celler som prolifererer raskt, eller for cellelinjer som har mistet evnen til å syntetisere ikke-essensielle aminosyrer de novo. Ved å lette den metabolske byrden ved biosyntesen bidrar det til forbedret vekstkinetikk, forlenget levedyktighet og større eksperimentell konsistens
- spesielt i næringssensitive kulturer eller kulturer med høy tetthet.
Sammensetning og bruk Løsningen inneholder glycin, L-alanin, L-asparagin, L-asparaginsyre, L-glutaminsyre, L-prolin og L-serin. Den er kompatibel med MEM og de fleste andre standardmedier. Fortynnes 1:100 i det endelige dyrkingsmediet. Dette produktet er sterilfiltrert og klart til bruk uten ytterligere håndteringstrinn. Kun til forskningsbruk. Ikke til bruk i diagnostiske eller terapeutiske prosedyrer. Ikke til bruk på mennesker eller dyr.

Accutase er en bruksklar, sterilt filtrert celleavløsningsløsning utviklet som et skånsomt alternativ til trypsin/EDTA for å løsne vedheftende pattedyrceller fra standard plastutstyr for vevskultur og overflater med vedheftbelegg. Den kombinerer proteolytisk og kollagenolytisk enzymaktivitet i en balansert saltløsning for å gi effektiv, men kontrollert løsning, samtidig som proteiner på celleoverflaten bevares og høy levedyktighet etter passering samt rask gjenfesting støttes.
Accutase-formuleringen er basert på Dulbeccos fosfatbufferte saltvann (DPBS) med EDTA og fenolrødt som en visuell pH-indikator. Enzymerne er av ikke-pattedyr
- og ikke-bakteriell opprinnelse, noe som gjør Accutase spesielt godt egnet til stamcelleforskning, vaksinearbeidsflyter og alle anvendelser der forurensninger av animalsk eller mikrobiell opprinnelse må minimeres. Løsningen hemmer seg selv ved 37 °C, så det kreves ikke noe nøytraliserende reagens eller serumholdig medium etter løsrivelse – cellene kan overføres direkte til ferskt medium.
Viktige egenskaper
Klar til bruk 1x sterilt filtrert væske – ingen fortynning eller rekonstituering nødvendig
Kombinert proteolytisk og kollagenolytisk enzymaktivitet for skånsom dissosiasjon
Hver batch er standardisert til en definert dissosiasjonsaktivitet for konsistens fra parti til parti
Enzymer av ikke-pattedyr
- og ikke-bakteriell opprinnelse
Selvhemmer ved 37 °C – ingen nøytraliserende løsning nødvendig
Formulert i Dulbeccos PBS med EDTA
Fenolrødt inkludert som visuell pH-indikator
pH 6,8–7,8
Typiske anvendelser
Accutase dissocierer skånsomt et bredt spekter av vedheftende og følsomme celletyper, inkludert humane embryonale stamceller (hESCs), humane induserte pluripotente stamceller (iPSCs), nevrale stamceller, primære nevroner og rutinemessig dyrkede vedheftende cellelinjer som HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 og A549. Typiske bruksområder inkluderer:
Rutinemessig subkultur og passering av vedheftende pattedyrceller
Skånsom enkeltcelle-dissociasjon av hESC, iPSC og andre følsomme linjer
Prøveforberedelse for strømningscytometri og FACS-analyse
Analyse av celleoverflatemarkører der epitopintegritet er viktig
Assayer for cellemigrasjon, proliferasjon og apoptose
Hvileassays ved serumdeprivasjon og onkogen-transfeksjonsstudier
Migrasjonstester av tumorceller og nevralkamceller
Oppskalering av produksjonen i bioreaktor-arbeidsflyter
For rutinearbeid, påfør ca. 10 ml Accutase per 75 cm2 kulturoverflate og inkuber i 5–10 minutter ved romtemperatur. Den optimale inkubasjonstiden bør bestemmes for hver cellelinje og bør ikke overstige en time. Før tilsetning, skyll cellelaget med en Ca2+/Mg2+-fri saltløsning, for eksempel DPBS uten kalsium og magnesium, for å fjerne rester av serum og toverdige kationer.
Håndtering og oppbevaring
Oppbevar den uåpnede flasken frosset ved -15 °C eller lavere. Tine enten ved romtemperatur eller over natten ved +2 °C til +8 °C. Ikke tine Accutase i et 37 °C vannbad, da høye temperaturer reduserer enzymaktiviteten. Etter tining kan løsningen oppbevares i opptil 2 måneder ved +2 °C til +8 °C; ikke oppbevar ved romtemperatur. Ikke forvarm reagensen til 37 °C før bruk – tilsett den direkte til vasket celler ved romtemperatur. For lang holdbarhet anbefales det å dele opp i engangsdoser for å unngå gjentatte tinesykluser. Arbeid alltid under aseptiske forhold.
Kvalitet
Produsert under strenge kvalitetsstandarder. Hver batch av Accutase er sterilt filtrert og testet for sterilitet, pH, utseende og dissosiasjonsaktivitet for å sikre konsistent, reproduserbar ytelse fra batch til batch.
Produktspesifikasjoner
Spesifikasjon
Detalj
ProdukttypeReagens for celleavløsning/dissosiasjon
FormatSterilfiltrert væske, klar til bruk
Volum100 ml
Arbeidskonsentrasjon1x (klar til bruk)
EnzymaktivitetKombinert proteolytisk og kollagenolytisk
Enzymets opprinnelseIkke-pattedyr og ikke-bakteriell
BuffersystemDulbeccos PBS med EDTA
pH-indikatorFenolrød
pH-område6,8 – 7,8
UtseendeKlar, lysrød til oransje løsning
Oppbevaringstemperatur-15 °C eller lavere
Stabilitet etter tiningOpptil 2 måneder ved +2 °C til +8 °C
Anbefalt bruksvolum~10 ml per 75 cm² kulturoverflate
Typisk inkubasjonstid5–10 minutter ved romtemperatur
FraktbetingelserFrosset på tørris
Tilsiktet brukKun til forskningsbruk og videre produksjon
Formulering (sammensetning per liter)
Komponent
Konsentrasjon (mg/L)
Uorganiske salter
Natriumklorid (NaCl)8000,00
Dinatriumhydrogenfosfat (Na2HPO4)1150,00
Kaliumklorid (KCl)200,00
Kaliumdihydrogenfosfat (KH2PO4)200,00
Andre komponenter
EDTA · 4Na (tetranatrium-EDTA)220,00
Fenolrød3,00
Proprietær enzymblanding (proteolytisk og kollagenolytisk aktivitet)1x
Accutase er et registrert varemerke tilhørende Innovative Cell Technologies, Inc.
, m: 2 mM L-Glutamin, m: 2,2 g/L NaHCO3, m: EBSS")
Denne bruksklare, sterilt filtrerte flytende formuleringen er tilsatt Earle’s Balanced Salt Solution (EBSS), 2 mM L-glutamin, D-glukose (1,0 g/l) og 2,2 g/l natriumbikarbonat (NaHCO3), noe som gjør den egnet for bruk i en CO2-kontrollert inkubatoratmosfære (typisk 5 % CO2). Det inkluderte fenolrød fungerer som en pH-indikator, noe som muliggjør praktisk visuell overvåking av mediumtilstanden under cellekulturen.
Viktige egenskaper
Klassisk Eagles MEM-formulering med Earles balanserte saltløsning (EBSS)
2 mM L-glutamin inkludert – klar til umiddelbar bruk
2,2 g/L natriumbikarbonat – bufret for inkubasjon med 5 % CO2
Med D-glukose (1,0 g/l) som primær karbonkilde
Med fenolrødt som pH-indikator
Uten HEPES og uten natriumpyruvat
Sterilfiltrert flytende medium, klar til bruk
pH 7,0 – 7,6
Typiske anvendelser
EMEM støtter dyrking av et bredt utvalg av pattedyrcellelinjer, inkludert HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 og mange primærceller. Vanlige bruksområder inkluderer:
Rutinemessig vedlikehold og utvidelse av vedheftende cellelinjer
Arbeidsflyter for virusformering og vaksineproduksjon
Cytotoksisitet og bioassay-anvendelser
Studier av transfeksjon og proteinekspresjon
Grunnleggende forskning innen cellebiologi og molekylærbiologi
For optimal cellevekst tilsettes EMEM vanligvis 5–10 % føtal bovint serum (FBS) og, avhengig av cellelinjen, ikke-essensielle aminosyrer (NEAA) og antibiotika som penicillin/streptomycin.
Håndtering og oppbevaring
Oppbevar den uåpnede flasken ved +2 °C til +8 °C, beskyttet mot lys. Etter åpning skal produktet brukes under aseptiske forhold. L-glutamin i løsning er utsatt for gradvis nedbrytning – vi anbefaler å bruke mediet innen 4 uker etter åpning for best ytelse, eller å tilsette fersk L-glutamin før bruk hvis det oppbevares i lengre perioder. La mediet varme seg opp til 37 °C før det tilsettes cellene.
Kvalitet
Produsert i henhold til strenge kvalitetsstandarder. Hver batch testes for sterilitet, pH, osmolalitet og endotoksinnivåer for å sikre jevn ytelse i cellekulturapplikasjoner.
Produktspesifikasjoner
Spesifikasjon
Detalj
ProdukttypeMEM
ProduktkategoriCellekulturmedier
FormatVæske
SteriltJa
Størrelse500 ml
L-glutaminMed L-glutamin (2 mM)
GlukoseMed glukose (1,0 g/l)
NatriumbikarbonatMed NaHCO3 (2,2 g/L)
HEPESUten HEPES
Natrium-pyruvatUten natriumpyruvat
FenolrødMed fenolrødt
SaltløsningEarle’s Balanced Salt Solution (EBSS)
pH7,0 – 7,6
EndotoksininnholdIkke spesifisert
Oppbevaring+2 °C til +8 °C
Formulering (sammensetning per liter)
Komponent
Konsentrasjon (mg/L)
Uorganiske salter
Kalsiumklorid · 2H2O265,00
Magnesiumsulfat97,72
Kaliumklorid400,00
Natriumklorid6 800,00
Natriumdihydrogenfosfat, vannfritt122,00
Natriumbikarbonat (NaHCO3)2 200,00
Aminosyrer
L-arginin · HCl126,00
L-cystin · 2HCl31,30
L-glutamin292,00
L-histidin · HCl · H2O42,00
L-isoleucin52,00
L-leucin52,00
L-lysin · HCl72,50
L-metionin15,00
L-fenylalanin32,00
L-treonin48,00
L-tryptofan10,00
L-tyrosin · 2Na · 2H2O51,90
L-valin46,00
Vitaminer
D-kalsiumpantotenat1,00
Kolin klorid1,00
Folsyre1,00
myo-inositol2,00
Nikotinamid1,00
Pyridoksal · HCl1,00
Riboflavin0,10
Tiamin · HCl1,00
Andre komponenter
D(+)-glukose1 000,00
Fenolrødt10,00

De viktigste egenskapene til Freeze Medium CM-1 inkluderer
Bred kompatibilitet: Effektivt for et bredt spekter av celletyper, inkludert primærceller, stamceller og etablerte cellelinjer.
Høy levedyktighet: Optimalisert for å maksimere cellegjenvinning og levedyktighet etter opptining, noe som sikrer pålitelige eksperimentelle resultater.
Klar til bruk: Praktisk tilberedt og sterilisert for umiddelbar bruk, noe som reduserer forberedelsestiden og risikoen for kontaminering.
Forbedret stabilitet: Opprettholder konsistent ytelse under standard kryopreserveringsforhold, noe som sikrer reproduserbare resultater.
Lang holdbarhet: CM-1 er et serumholdig kryopreserveringsmedium som er klart til bruk, og som kan oppbevares i kjøleskapet i opptil ett år.
Bruk av CM-1 til nedfrysing av celler
Følg disse trinnene for å bruke CM-1 til nedfrysing av både adherente celler og suspensjonsceller
For adherente celler, vask og dissosier dem fra kultursubstratet. For suspensjonsceller går du direkte til neste trinn.
Tell cellene for å sikre at de har riktig konsentrasjon.
Sentrifuger cellene for å pelleter dem, og resuspender dem deretter i CM-1-frysemedium.
Overfør de resuspenderte cellene til kryovialer.
Bruk en langsom nedfrysingsmetode før du overfører cellene til langtidslagring
Metode
Beskrivelse
Trinn
❄️
Manuell frysing
En trinnvis metode som innebærer gradvis reduksjon av temperaturen for å sikre cellenes levedyktighet
1️⃣ Plasser celler i frysemedium i en fryser på 4 °C i 40 minutter.
2️⃣ Overfør til en fryser på -80 °C i 24 timer.
3️⃣ Oppbevar cellene i flytende nitrogen for langtidsoppbevaring
❄️
Bruk av Mr. Frosty
Et praktisk apparat som gjør det mulig å kontrollere frysehastigheten uten elektrisk strøm
1️⃣ Klargjør celler i kryovialer med frysemedium.
2️⃣ Plasser kryovialene i Mr. Frosty-beholderen.
3️⃣ Oppbevares ved -80 °C i 24 timer før de overføres til flytende nitrogen
❄️
Fryser med kontrollert hastighet
En høypresisjonsfryser fra Thermo Fisher eller andre produsenter som er utviklet for kontrollert temperaturreduksjon
1️⃣ Programmer enheten til å senke temperaturen gradvis.
2️⃣ Plasser de forberedte cellene i fryseren.
3️⃣ Etter frysesyklusen overføres cellene til flytende nitrogen
Oppbevar kryovialene ved temperaturer under -130 °C eller i flytende nitrogen for langtidsoppbevaring.
Ingredienser
Inneholder FBS, DMSO, glukose, salter
Bufferkapasitet: pH = 7,2 til 7,6
Cytions frysemedium CM-1 er en pålitelig løsning for kryopreservering som sikrer høy cellelevedyktighet og funksjonalitet etter opptining for et bredt spekter av forskningsapplikasjoner.

- og kyllingleverceller, spesielt under forhold med redusert serum.
Ham's F-12K (Kaighn's) Medium er nøye sammensatt for å optimalisere forholdene for celledyrking. Det har en beriket sammensetning som gir forhøyede nivåer av essensielle komponenter som aminosyrer og natriumpyruvat, samt tilleggselementer som putrescin, tymidin, hypoksantin og sink. Disse tilsetningene gjør det mulig for forskere å supplere mediet med minimalt med serum eller definerte komponenter for spesifikke celletyper, noe som forenkler presise eksperimentelle forhold.
Ham's F-12K (Kaighn's) Medium inneholder ikke proteiner eller vekstfaktorer. Derfor er det ofte nødvendig å supplere med vekstfaktorer og føtalt bovint serum (FBS), slik at forskerne kan skreddersy mediet etter kravene til sine spesifikke cellelinjer. For å oppnå optimal ytelse må konsentrasjonen av FBS optimaliseres nøye for hver enkelt cellelinje, slik at optimal vekst og funksjonalitet sikres.
For å opprettholde en fysiologisk pH benytter Ham's F-12K (Kaighn's) Medium et natriumbikarbonatbuffersystem (2,5 g/L), noe som krever et kontrollert CO2-miljø på 5-10 % under dyrkingen. Dette sikrer at pH-verdien i mediet holder seg innenfor det ideelle området for cellevekst og levedyktighet
Kvalitetskontroll
pH = 7,2 +/
- 0,02 ved 20-25 °C.
Hvert parti er testet for sterilitet og fravær av mykoplasma og bakterier.
Vedlikehold
Oppbevares i kjøleskap ved +2 °C til +8 °C i mørke. Frysing og oppvarming opp til +37 °C reduserer produktets kvalitet.
Ikke varm opp mediet til mer enn 37 °C eller bruk ukontrollerbare varmekilder (f.eks. mikrobølgeovner).
Hvis bare en del av mediet skal brukes, tar du denne mengden ut av flasken og varmer den opp i romtemperatur.
Holdbarhetstiden for alle medier unntatt basismediet er 8 uker fra produksjonsdato.
Sammensetning
Komponenter
mg/L
Uorganiske salter
Kalsiumklorid x 2H2O
135,24
Kobber(II)sulfat x 5H2O
0,00
Jern(II)sulfat x 7H2O
0,83
Magnesiumklorid x 6H2O
105,72
Magnesiumsulfat x 7H2O
394,49
Kaliumklorid
283,29
Kaliumdihydrogenfosfat
58,52
Natriumklorid
7597,20
di-Natriumhydrogenfosfatvannfritt
115,02
Sink sulfat x 7H2O
0,14
Andre komponenter
D(+)-Glukose vannfri
1260,00
Hypoksantin
4,08
DL-α-Liponsyre
0,21
Fenolrød
3,00
Putrescin x 2HCl
0,32
Natriumpyruvat
220,00
NaHCO3
2500,00
Tymidin
0,73
Aminosyrer
L-Alanin
17,82
L-arginin x HCl
421,40
L-asparagin x H2O
30,02
L-asparaginsyre
26,62
L-Cystein x HCl x H2O
70,24
L-glutamin
292,20
L-glutaminsyre
29,42
Glysin
15,01
L-Histidin x HCl x H2O
41,92
L-isoleucin
7,87
L-Leucin
26,24
L-Lysin x HCl
73,04
L-metionin
8,95
L-fenylalanin
9,91
L-prolin
69,06
L-Serin
21,02
L-Threonin
23,82
L-Tryptofan
4,08
L-Tyrosin
10,87
L-Valin
23,42
Vitaminer
D(+)-Biotin
0,07
D-kalsiumpantotenat
0,48
Kolinklorid
13,96
Folsyre
1,32
myo-Inositol
18,02
Nikotinamid
0,04
Pyridoksin x HCl
0,06
Riboflavin
0,04
Tiamin x HCl
0,34
Vitamin B12
1,36

Fosfatbufret saltvann (PBS) er en mye brukt bufferløsning i biologisk og kjemisk forskning. Den spiller en avgjørende rolle når det gjelder å opprettholde pH-balansen og osmolariteten under ulike eksperimentelle prosedyrer, inkludert vevsprosessering og cellekultur. Vår PBS-løsning er omhyggelig formulert med ingredienser av høy renhet for å sikre stabilitet og pålitelighet i hvert eneste eksperiment. Osmolariteten og ionekonsentrasjonene i PBS-løsningen vår etterligner i stor grad de som finnes i menneskekroppen, noe som gjør den isotonisk og ugiftig for de fleste celler.
Sammensetningen av vår PBS-løsning
Vår PBS-løsning er en pH-justert blanding av ultrarene fosfatbuffere og saltvannsløsninger. Ved en 1X arbeidskonsentrasjon inneholder den
8000 mg/L natriumklorid (NaCl)
200 mg/L kaliumklorid (KCl)
1150 mg/L vannfritt dibasisk natriumfosfat (Na2HPO4)
200 mg/L Vannfritt kaliumfosfat monobasisk (KH2PO4)
Denne sammensetningen sikrer en optimal pH
- og ionebalanse, og er egnet for et bredt spekter av biologiske anvendelser.
Bruksområder for vår PBS-løsning
Vår PBS-løsning er ideell for ulike bruksområder innen biologisk forskning. De isotone og giftfrie egenskapene gjør den egnet til fortynning av substanser og skylling av cellebeholdere. PBS-løsninger som inneholder EDTA, er effektive for å løsne celler som sitter fast og klumper seg sammen. PBS bør imidlertid ikke tilsettes toverdige metaller som sink, da dette kan føre til utfelling. I slike tilfeller anbefales Good's buffere. I tillegg er vår PBS-løsning et akseptabelt alternativ til viralt transportmedium for transport og lagring av RNA-virus, inkludert SARS-CoV-2.
Kvalitetskontroll
Sterilfiltrert
Oppbevaring og holdbarhet
Oppbevares ved +2 °C til +25 °C, beskyttet mot lys.
Etter åpning, oppbevares ved 2 °C til 25 °C og brukes innen 24 måneder.
Transportforhold
Omgivelsestemperatur
Vedlikehold
Oppbevares i kjøleskap ved +2 °C til +8 °C i mørke. Unngå frysing og hyppig oppvarming til +37 °C, da det reduserer produktkvaliteten.
Ikke varm opp mediet til mer enn 37 °C eller bruk ukontrollerte varmekilder som mikrobølgeovner.
Hvis bare en del av mediet skal brukes, ta ut den nødvendige mengden og varm den opp til romtemperatur før bruk.
Sammensetning
Kategori
Komponenter
Konsentrasjon (mg/L)
Salter
Kaliumklorid
200
Kaliumfosfat, vannfritt, monobasisk
200
Natriumklorid
8000
Natriumfosfat, vannfritt, dibasisk
1150

RPMI 1640 Medium ble opprinnelig utviklet for å støtte veksten av humane leukemiceller i både suspensjons
- og monolagskulturer, men har gjennom modifikasjoner fra forskere og kommersielle leverandører utviklet seg til å bli egnet for et bredt spekter av pattedyrceller. Det er eksepsjonelt kompatibelt med cellelinjer som HeLa, Jurkat, MCF-7, PC12, PBMC, astrocytter og karsinomer.
RPMI 1640 Medium skiller seg ut fra andre cellekulturmedier på grunn av sin unike sammensetning. Det inneholder en betydelig mengde fosfat, aminosyrer og vitaminer. Det inneholder blant annet biotin, vitamin B12 og PABA, som ikke finnes i Eagle's Minimal Essential Medium eller Dulbecco's Modified Eagle Medium. RPMI 1640 Medium har dessuten betydelig høyere konsentrasjoner av vitaminene inositol og kolin. Det inneholder imidlertid ikke proteiner, lipider eller vekstfaktorer. Derfor er det ofte nødvendig å tilsette 10 % føtalt bovint serum (FBS) for å skape optimale forhold for cellevekst.
Buffersystemet i RPMI 1640 Medium er basert på natriumbikarbonat og krever et 5-10 % CO2-miljø for å opprettholde en fysiologisk passende pH. Inkluderingen av reduksjonsmidlet glutation skiller dette mediet ytterligere fra andre.
Kvalitetskontroll
Sterilfiltrert
Oppbevaring og holdbarhet
Oppbevares ved +2 °C til +8 °C, beskyttet mot lys.
Etter åpning, oppbevar ved 4 °C og bruk innen 6-8 uker.
Transportforhold
Omgivelsestemperatur
Vedlikehold
Oppbevares i kjøleskap ved +2 °C til +8 °C i mørke. Unngå frysing og hyppig oppvarming til +37 °C, da det reduserer produktkvaliteten.
Ikke varm opp mediet til mer enn 37 °C eller bruk ukontrollerte varmekilder som mikrobølgeovner.
Hvis bare en del av mediet skal brukes, ta ut den nødvendige mengden og varm den opp til romtemperatur før bruk.
Sammensetning
Kategori
Komponenter
Konsentrasjon (mg/L)
Aminosyrer
Glysin
10.00
L-Alanyl-L-glutamin
434.40
L-arginin
200.00
L-asparaginH2O
56.82
L-asparaginsyre
20.00
L-Cystin 2HCl
65.20
L-glutaminsyre
20.00
L-Histidin HClH2O
20.27
L-hydroksy-L-prolin
20.00
L-isoleucin
50.00
L-Leucin
50.00
L-Lysin HCl
40.00
L-metionin
15.00
L-fenylalanin
15.00
L-prolin
20.00
L-Serin
30.00
L-Threonin
20.00
L-Tryptofan
5.00
L-Tyrosin 2Na 2H2O
28.83
L-Valin
20.00
Vitaminer
p-aminobensoesyre
1.00
D-Biotin
0.20
Kolinklorid
3.00
D-kalsiumpantotenat
0.25
Folsyre
1.00
myo-Inositol
35.00
Nikotinamid
1.00
Pyridoksin HCl
1.00
Riboflavin
0.20
Tiamin HCl
1.00
Vitamin B12
0.005
Uorganiske salter
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Andre komponenter
D-Glukose
2000.00
L-glutation redusert
1.00
Fenolrødt natriumsalt
5.30
, w: 3,1 g/L glukose, w: 2,5 mM L-glutamin, w: 15 mM HEPES, w: 0,5 mM natriumpyruvat, w: 1,2 g/L NaHCO3")
Denne unike formuleringen kombinerer Dulbeccos Modified Eagle Medium (DMEM) og Ham's F-12 (Ham's Nutrient Mixture F-12) i et nøyaktig 1:1-forhold. Tilsetningen av L-glutamin forbedrer sammensetningen ytterligere.
DMEM, avledet fra Eagles Minimal Essential Medium (EMEM), tilbyr en økt konsentrasjon av aminosyrer og vitaminer sammenlignet med sin forgjenger. I kontrast er Hams F-12 basert på Hams F-10-medium, som gir et komplementært sett av essensielle komponenter.
For å støtte optimal cellevekst er det vanlig praksis å supplere DMEM:Ham's F12 med FBS i en typisk konsentrasjon på 5–10 %. Dette tilskuddet er nødvendig, da mediet mangler veksthormoner, lipider og proteiner som er avgjørende for celleutviklingen.
DMEM:Ham's F12 inneholder et pH-buffersystem og tilsettes ofte fenolrødt, en pH-indikator. Dyrkede celler i DMEM:Ham's F12, eller ethvert medium som bruker bikarbonatbuffersystemet, krever et kontrollert CO2-miljø på 5–10 % for å opprettholde riktige pH-nivåer.
Kvalitetskontroll
Sterilfiltrert
Oppbevaring og holdbarhet
Oppbevares ved +2 °C til +8 °C, beskyttet mot lys.
Etter åpning skal produktet oppbevares ved 4 °C og brukes innen 6–8 uker.
Fraktbetingelser
Omgivelsestemperatur
Oppbevaring
Oppbevares i kjøleskap ved +2 °C til +8 °C i mørke. Unngå frysing og hyppig oppvarming til +37 °C, da dette reduserer produktkvaliteten.
Ikke varm opp mediet til over 37 °C eller bruk ukontrollerte varmekilder som mikrobølgeovner.
Hvis bare en del av mediet skal brukes, fjern den nødvendige mengden og varm den opp til romtemperatur før bruk.
Sammensetning
Kategori
Komponenter
Konsentrasjon (mg/L)
Aminosyrer
Glycin
18,75
L-alanin
4,45
L-arginin-HCl
147,50
L-asparagin H₂O
7,50
L-asparaginsyre
6,65
L-cystein HCl H₂O
17,56
L-cystin 2 HCl
31,29
L-glutaminsyre
7,35
L-glutamin
365,00
L-histidin-HCl-H₂O
31,48
L-isoleucin
54,47
L-leucin
59,05
L-lysin-HCl
91,25
L-metionin
17,24
L-fenylalanin
35,48
L-prolin
17,25
L-serin
26,25
L-treonin
53,45
L-tryptofan
9,02
L-tyrosin 2 Na 2 H2O
55,79
L-valin
52,85
Vitaminer
D-biotin
0,0035
Kolin klorid
8,98
D-kalsiumpantotenat
2,24
Folsyre
2,66
myo-inositol
12,60
Nikotinamid
2,02
Pyridoksin-HCl
0,031
Pyridoksal-HCl
2,00
Riboflavin
0,219
Tiamin-HCl
2,17
Vitamin B12
0,68
Uorganiske salter
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO3)3 9 H2O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Andre komponenter
D-glukose
3151,00
Hypoxantin
2,40
HEPES
3574,50
Linolsyre
0,042
Liponsyre
0,105
Fenolrødt natriumsalt
8,63
Putrescin-2-HCl
0,081
Natrium-pyruvat
55,00
Tymidin
0,365
, m: 3,0 g/L glukose, m: stabil glutamin, m: 2,0 mM natriumpyruvat, m: 2,2 g/L NaHCO3")
- 0,02 ved 20-25 °C. Hvert parti er testet for sterilitet og fravær av mykoplasma og bakterier. Oppbevaring Oppbevares i kjøleskap ved +2 °C til +8 °C i mørket. Frysing og oppvarming til +37 °C reduserer produktets kvalitet. Ikke varm opp mediet til over 37 °C og ikke bruk ukontrollerbare varmekilder (f.eks. mikrobølgeovn). Hvis bare en del av mediet skal brukes, fjern denne mengden fra flasken og varm den opp til romtemperatur. Holdbarheten for alle medier unntatt basismediet er 6 til 8 uker fra åpningsdatoen. Sammensetning Komponenter mg/L Uorganiske salterKalsiumklorid x 2H2O132,00 Magnesiumsulfat97,67 Kaliumklorid400 Natriumklorid6 460,00 Dinatriumhydrogenfosfat (vannfritt)504,00 Andre komponenterD(+)-glukose (vannfri)3 000 Glutathion (redusert)0,5 Kjøttpeptone600 Fenolrødt natriumsalt11 AminosyrerL-alanin13,36 L-arginin x HCl42,14 L-asparagin x H2O45,03 L-asparaginsyre19,97 L-Cystein x HCl x H2O31,75 L-glutamin (stabil)219,15 L-glutaminsyre22,07 Glysin7,51 L-histidin x HCl x H2O20,96 L-hydroksyprolin19,67 L-isoleucin39,36 L-leucin39,36 L-lysin x HCl36,54 L-metionin14,92 L-fenylalanin16,52 L-prolin17,27 L-serin26,28 L-treonin17,87 L-tryptofan3,06 L-tyrosin dinatriumsalt26,1 L-valin17,57 Vitaminerp-Aminobensoesyre1,0 Askorbinsyre0,56 D(+)-biotin0,2 D-kalsiumpantotenat0,2 Kolin klorid5 Folsyre10 Myo-inositol36 Nikotinamid0,5 Nikotinsyre0,5 Pyridoksal-HCl0,5 Pyridoksin HCl0,5 Riboflavin0,2 Tiamin HCl0,2 Vitamin B122,0

Medium 199 har en rekke bruksområder innen feltet. Det kan effektivt opprettholde kumulus-oocytt-komplekset (COC) og støtte in vitro-modning av oocytter. I tillegg brukes det til skylling av aspirasjonslinjer under egginnsamling fra tyske Holstein-kyr. Medium 199 er dessuten et utmerket medium for dyrking av hjerteendotelceller fra rotter. Disse bruksområdene viser hvor allsidig og tilpasningsdyktig Medium 199 er til ulike eksperimentelle behov.
Historikk
Utviklingen av Medium 199 på 1950-tallet markerte et betydelig fremskritt innen vevskulturmedier. Før det ble introdusert, var mange dyrkingsmedier basert på produkter og vevsekstrakter fra dyr. Morgan og hans kolleger revolusjonerte imidlertid feltet ved å formulere en fullstendig definert næringskilde for cellekulturer. Gjennom eksperimenter med ulike kombinasjoner av vitaminer, aminosyrer og andre faktorer oppdaget de de eksepsjonelle vekstfremmende egenskapene til Medium 199.
Kvalitetskontroll
pH = 7,2 +/
- 0,02 ved 20-25 °C.
Hvert parti er testet for sterilitet og fravær av mykoplasma og bakterier.
Vedlikehold
Oppbevares i kjøleskap ved +2 °C til +8 °C i mørke. Frysing og oppvarming opp til +37 °C minimerer produktets kvalitet.
Ikke varm opp mediet til mer enn 37 °C eller bruk ukontrollerbare varmekilder (f.eks. mikrobølgeovner).
Hvis bare en del av mediet skal brukes, tar du denne mengden ut av flasken og varmer den opp i romtemperatur.
Holdbarheten for alle medier unntatt basismediet er 8 uker fra produksjonsdatoen.
Sammensetning
Komponenter
mg/L
Uorganiske salter
Kalsiumklorid x 2H2O
264,92
Jern(III)nitrat x 9H2O
0,72
Magnesiumsulfat
97,67
Kaliumklorid
400,00
Natriumacetat x 3H2O
82,95
Natriumklorid
6,800.00
Natriumdihydrogenfosfat x H2O
140,00
Andre komponenter
Adeninsulfat
10,00
AMP
0,20
ATP
1,00
Kolesterol
0,20
2'-Deoksyribose
0,50
D(+)-Glukose vannfri
1,000.00
Glutation (rød)
0,05
Guanin x HCl
0,30
Hypoksantin
0,30
Fenol rød
10,00
D-Ribose
0,50
Tymin
0,30
Tween 80
4,90
Uracil
0,30
Xantin
0,30
NaHCO3
2,200.00
Aminosyrer
L-Alanin
25,00
L-arginin x HCl
70,00
L-asparaginsyre
30,00
L-Cystein x HCl x H2O
0,10
L-Cystin
20,00
L-glutamin stabil
149,00
L-glutaminsyre
67,00
Glysin
50,00
L-Histidin x HCl x H2O
21,88
L-Hydroksyprolin
10,00
L-isoleucin
20,00
L-Leucin
60,00
L-Lysin x HCl
70,00
L-metionin
15,00
L-fenylalanin
25,00
L-prolin
40,00
L-Serin
25,00
L-Threonin
30,00
L-Tryptofan
10,00
L-Tyrosin
40,00
L-Valin
25,00
Vitaminer
4-Amino benzosyre
0,05
Askorbinsyre
0,05
D(+)-Biotin
0,01
Calciferol
0,10
D-kalsiumpantotenat
0,01
Kolinklorid
0,50
Folsyre
0,01
myo-Inositol
0,05
Menadion
0,01
Nikotinsyre
0.025
Nikotinamid
0.025
Pyridoksal x HCl
0.025
Pyridoksol x HCl
0.025
Riboflavin
0,01
DL-α-tokoferolfosfat dinatriumsalt
0,01
Tiamin x HCl
0,01
Vitamin A-acetat
0,14

IMDM egner seg godt til raskt prolifererende cellekulturer med høy tetthet, inkludert Jurkat-, COS-7
- og makrofagceller. De ulike modifikasjonene av IMDM som er tilgjengelige for en rekke ulike cellekulturapplikasjoner, finner du ved hjelp av medievælgerverktøyet. Flytende medier gir viktige næringsstoffer til alle celledyrkingsapplikasjoner. Alle våre høykvalitetsmedier for celledyrking produseres i henhold til den opprinnelig publiserte formelen eller de modifikasjonene som er nødvendige for å sikre konsekvent ytelse og stabilitet for dyrkingsmediet.
IMDM vs. DMEM
IMDM inneholder kaliumnitrat i stedet for jernnitrat og HEPES og natriumpyruvat. De ekstra komponentene i IMDM gjør det mer egnet for spesialiserte celletyper og spesifikke bruksområder enn DMEM.
IMDM vs. RPMI
IMDM og RPMI har forskjellige formuleringer som kan være relevante for PMA/ionomycin-stimulering. En vesentlig forskjell er konsentrasjonen av Ca2+. Mens RPMI inneholder 0,42 mM Ca2+, inneholder IMDM 1,49 mM.
Kvalitetskontroll
pH = 7,2 +/
- 0,02 ved 20-25 °C.
Hvert parti er testet for sterilitet og fravær av mykoplasma og bakterier.
Vedlikehold
Oppbevares i kjøleskap ved +2 °C til +8 °C i mørke. Frysing og oppvarming opp til +37 °C minimerer produktets kvalitet.
Ikke varm opp mediet til mer enn 37 °C eller bruk ukontrollerbare varmekilder (f.eks. mikrobølgeovner).
Hvis bare en del av mediet skal brukes, tar du denne mengden ut av flasken og varmer den opp i romtemperatur.
Holdbarheten for alle medier unntatt basismediet er 8 uker fra produksjonsdatoen.
Sammensetning
Komponenter
mg/L
Uorganiske salter
Kalsiumklorid x 2 H2O
219,00
Kaliumklorid
330,00
Kaliumnitrat
0.076
Magnesiumsulfat vannfritt
97,73
Natriumklorid
4,505.00
Natriumdihydrogenfosfat vannfritt
109,00
Natriumselenitt
0,02
Andre komponenter
D(+)-Glukose vannfri
4,500.00
HEPES
5,958.00
Natriumpyruvat
110,00
Fenolrød
15,00
Aminosyrer
L-Alanin
25,00
L-arginin x HCl
84,00
L-asparagin x H2O
25,00
L-asparaginsyre
30,00
L-Cystin x 2HCl
91,24
L-glutamin
584,00
L-glutaminsyre
75,00
Glysin
30,00
L-Histidin x HCl x H2O
42,00
L-isoleucin
104,80
L-Leucin
104,80
L-Lysin x HCl
146,20
L-metionin
30,00
L-fenylalanin
66,00
L-prolin
40,00
L-Serin
42,00
L-Threonin
95,20
L-Tryptofan
16,00
L-Tyrosin x 2Na
104,20
L-Valin
93,60
Vitaminer
D(+)-Biotin
0.013
D-kalsiumpantotenat
4,00
Kolinklorid
4,00
Folsyre
4,00
myo-Inositol
7,20
Cellekulturmedier: En oversikt
Innenfor biovitenskapene er cellekultur en av de viktigste metodene. Begrepet «cellekultur» refererer til uttak av celler, vev eller organer fra et dyr eller en plante og påfølgende implantering av disse cellene, vevene eller organene i et kunstig miljø som er gunstig for deres overlevelse og/eller vekst. De grunnleggende miljøkravene for optimal celleutvikling er kontrollert temperatur, et underlag for cellefesting, et passende vekstmedium og en inkubator som opprettholder optimal pH og osmolalitet. Cellene må ha disse forholdene for å kunne vokse til sitt fulle potensial.
Valget av et egnet vekstmedium for in vitro-dyrking er den fasen i cellekulturen som er både den mest kritiske og den viktigste. Et vekstmedium, også kjent som dyrkningsmedium, er en væske eller gel som er sammensatt for å fremme organismers utvikling på mikroskopisk, cellulært eller planteaktig nivå. Mediet som brukes til å dyrke celler inneholder ofte en tilstrekkelig tilførsel av energi og stoffer som regulerer cellesyklusen. Hovedkomponentene i et dyrkningsmedium inkluderer aminosyrer, vitaminer, uorganiske salter, glukose og serum. Serumet tilsettes mediet fordi det fungerer som en kilde til vekstfaktorer, hormoner og vedheftsfaktorer. I tillegg til å tilføre næringsstoffer, bidrar mediet også til å opprettholde pH- og osmolalitetsnivåene.
Typer av medier som brukes i cellekultur
Både menneskelige og dyreceller kan dyrkes enten i et kunstig eller syntetisk medium eller i et helt naturlig medium som er tilsatt naturlige elementer. I det følgende gir vi deg en oversikt over de ulike medietyper som er tilgjengelige i dag.
Naturlige medier
Naturlige medier inneholder kun biologiske væsker som finnes i sin naturlige tilstand. Naturlige medier er svært nyttige og enkle å bruke for dyrking av et bredt spekter av dyrecelletyper. Manglende forståelse av de nøyaktige komponentene som utgjør naturlige medier, er den viktigste faktoren som bidrar til den lave repeterbarheten av resultater oppnådd ved bruk av naturlige medier.
Kunstige medier
Fremstillingen av kunstige eller syntetiske medier innebærer tilsetning av næringsstoffer (både organiske og uorganiske), serumproteiner, karbohydrater, kofaktorer, vitaminer og salter, samt O₂- og CO₂-gassfaser [1].
Det er utviklet ulike typer kunstige medier for å oppfylle en eller flere av følgende funksjoner: 1) Umiddelbar overlevelse (en balansert saltløsning med nøyaktig pH og osmotisk trykk). 2) Langvarig overlevelse (en balansert saltløsning tilsatt ulike formuleringer av organiske kjemikalier og/eller serum). 3) Ubegrenset utvikling. 4) Spesialiserte funksjoner.
Det finnes fire forskjellige klassifiseringer av kunstige medier:
Serumholdige medier
Den vanligste typen tilsetning i medier som brukes til dyrking av dyreceller er føtalt bovint serum. Det tilsettes dyrkingsmediet som et kostnadseffektivt tilskudd for å oppnå best mulige vekstforhold. I tillegg til å fungere som transportør eller chelator for næringsstoffer som er ustabile eller vannuoppløselige, hormoner og vekstfaktorer, proteasehemmere og andre stoffer, binder og nøytraliserer serumet også skadelige molekyler.
Serumfritt medium
Tilstedeværelsen av serum i mediet har en rekke ulemper og kan potensielt føre til alvorlige tolkningsfeil i immunologisk forskning [2, 3]. Det er utviklet en rekke ulike serumfrie medier [4, 5]. Disse mediene er generelt spesielt formulert for å støtte dyrking av én enkelt celletype, slik som Knockout Serum Replacement og Knockout DMEM fra Thermo Fisher Scientific, samt mTESR-mediet fra Stem Cell Technologies [6], beregnet på stamceller [7].
I tillegg inneholder disse mediene definerte mengder rensede vekstfaktorer, lipoproteiner og andre proteiner, som ellers vanligvis tilføres via serumet [8]. Disse mediene kalles ofte «definerte dyrkingsmedier», siden komponentene som inngår i dem er godt kjent.
Kjemisk definerte medier
Disse mediene inneholder ultrarene uorganiske og organiske komponenter som ikke er forurenset av noen form for kontaminasjon. De kan også inneholde tilsetninger av rene proteiner, for eksempel vekstfaktorer.
Genmodifisering av bakterier eller gjær, sammen med tilsetning av bestemte fettsyrer, vitaminer, kolesterol og aminosyrer, resulterer i produksjonen av deres bestanddeler [9].
Proteinfri næringsmedier
Proteinfri næringsmedier er de som ikke inneholder noe protein i det hele tatt, men i stedet kun består av ikke-proteinholdige elementer. Sammenlignet med næringsmedier med tilsatt serum fremmer bruk av næringsmedier uten tilsatt protein større celleproliferasjon og proteinekspresjon, og gjør det enklere å rense ethvert produkt som genereres i en nedstrømsprosess [10–12]. Protein inngår ikke i formuleringer som MEM og RPMI-1640. Imidlertid kan et proteintilskudd tilsettes dersom det er nødvendig.
Dyrkningsmedier og deres grunnleggende komponenter
Kommersielle dyrkningsmedier kan kjøpes som pulver eller væske og inneholder ofte en rekke næringsstoffer som aminosyrer, glukose, salter, vitaminer og andre kosttilskudd.
Behovet for disse komponentene varierer fra cellelinje til cellelinje, og disse variasjonene er årsaken til det store antallet ulike formuleringer av medier. Hver komponent har en bestemt funksjon, som vil bli beskrevet i de følgende avsnittene:
Buffersystemer
For å opprettholde optimale vekstforhold må pH-verdien kontrolleres, noe som ofte gjøres ved hjelp av ett av to buffersystemer:
Naturlig buffersystem
Forholdet mellom CO₂ og H₂CO₃ i atmosfæren er det samme som i mediet, noe som skaper en naturlig buffermekanisme. For å bevare denne naturlige buffermekanismen må kulturene holdes i et luftmiljø med 5–10 % CO₂, noe som ofte oppnås ved bruk av en CO₂-inkubator. En av de største fordelene ved å bruke en naturlig buffer er at den er billig og sikker.
HEPES
Kjemisk buffering ved hjelp av zwitterionen HEPES har større bufferevne i pH-området 7,2–7,4 og krever ikke et regulert gassmiljø. For visse celletyper kan en større dose HEPES være skadelig. Medier som inneholder HEPES er også mye mer utsatt for de fototoksiske effektene av fluorescerende lys [13].
Fenolrødt
pH-indikatoren fenolrød inngår ofte i kommersielt tilgjengelige dyrkningsmedier, noe som muliggjør kontinuerlig overvåking av pH. Når cellene vokser, forårsaker metabolittene som produseres av disse cellene en endring i pH og dermed en fargeendring i mediet. Fenolrød har en dobbel effekt på mediets farge: det blir gult ved surt pH og lilla ved alkalisk pH. pH 7,4, den optimale verdien for cellekultur, gjør at mediet fremstår som fluorescerende rødt.
Men fenolrød har noen ulemper: For det første kan fenolrød etterligne virkningen av en rekke steroidhormoner, først og fremst østrogen [14]. Derfor anbefales det å bruke et medium uten fenolrød når man studerer østrogenfølsomme celler som brystvev. Natrium-kaliumbalansen forstyrres av tilstedeværelsen av fenolrødt i flere serumfrie formuleringer. Tilsetning av serum eller hypofysehormon fra storfe til mediet kan motvirke denne effekten [15]. For det tredje hindres deteksjonen i strømningscytometriske eksperimenter av tilstedeværelsen av fenolrødt.
Uorganiske salter
Medier som inneholder uorganiske salter, som natrium-, kalium- og kalsiumioner, bidrar til å opprettholde osmotisk likevekt og regulere membranpotensialet.
Aminosyrer
Siden aminosyrer er de grunnleggende komponentene i protein, er de en essensiell del av hvert eneste cellevekstmedium som noensinne er utviklet. Fordi celler ikke er i stand til å produsere visse aminosyrer på egen hånd, er det viktig at dyrkningsmediet inneholder essensielle aminosyrer. De er nødvendige for celleproliferasjon, og konsentrasjonen de forekommer i bestemmer den maksimale celletettheten som kan oppnås. Spesielt er L-glutamin, en essensiell aminosyre, av avgjørende betydning.
L-glutamin fungerer som en sekundær energikilde for stoffskiftet og bidrar med nitrogen til produksjonen av NAD, NADPH og nukleotider. Siden L-glutamin er en ustabil aminosyre som over tid omdannes til en form som cellene ikke kan utnytte, må den tilsettes mediet.
I tillegg kan ikke-essensielle aminosyrer tilsettes vekstmediet for å erstatte de som er blitt forbrukt under vekstprosessen. Cellenes vekst stimuleres og levedyktigheten økes når vekstmediet tilsettes ikke-essensielle aminosyrer.
Karbohydrater
Karbohydrater i form av sukker er den viktigste energikilden. Mange av mediene inneholder også maltose og fruktose i tillegg til de mer vanlige sukkerartene glukose og galaktose.
Proteiner og peptider
Albumin, transferrin og fibronektin er de mest brukte proteinene og peptidene. De er spesielt viktige i medier som ikke inneholder serum. Albumin, transferrin, aprotinin, fetuin og fibronektin er noen av proteinene som kan finnes i serum, som er en rik proteinkilde.
Albumin er det viktigste proteinet i blodet, og dets funksjon er å binde og transportere ulike stoffer, blant annet vann, salter, frie fettsyrer, hormoner og vitaminer, mellom ulike organer og celler. Albumins evne til å binde seg til kjemikalier gjør det til et effektivt middel for å fjerne skadelige forbindelser fra mediet der cellene dyrkes.
Aprotinin er et beskyttende middel i cellekultursystemer, siden det er stabilt ved nøytral og sur pH, samt motstandsdyktig mot høye temperaturer og nedbrytning som kan forårsakes av proteolytiske enzymer. Det er i stand til å hemme en rekke serinproteaser, blant annet trypsin.
Fetuin er et glykoprotein som kan påvises i større mengder i serumet til foster og nyfødte dyr sammenlignet med serum fra voksne. I tillegg fungerer det som en serinproteasehemmer. Proteinet fibronektin er en essensiell komponent i prosessen med celleadhesjon. Transferrin er et protein som transporterer jern og er ansvarlig for å levere jern til cellemembranene.
Fettsyrer og lipider
De spiller en avgjørende rolle i serumfritt medium når serum mangler.
Vitaminer
Mange vitaminer er nødvendige for cellenes utvikling og proliferasjon. Vitaminer kan ikke produseres i tilstrekkelige mengder av cellene og er derfor essensielle i vevskultur som kosttilskudd.
I cellekultur er serum den primære kilden til vitaminer; imidlertid tilsettes også ulike vitaminer i mediet for å tilpasse det til en bestemt celletype. Vanligvis brukes B-vitaminer for å stimulere veksten.
Sporstoffer
Kjemiske elementer som kobber, sink, selen og trikarboxylsyre-mellomprodukter kalles sporstoffer. Sporstoffer tilsettes ofte i medier som ikke inneholder serum for å erstatte de som vanligvis finnes i serum. Disse elementene er viktige kjemiske komponenter som er nødvendige for en sunn celleutvikling. Mange biokjemiske reaksjoner er avhengige av visse mikronæringsstoffer, for eksempel enzymaktivitet.
Medietilskudd
Det komplette vekstmediet som anbefales for visse cellelinjer, trenger ekstra komponenter som ikke finnes i basismediet og serumet. Disse kosttilskuddene støtter cellevekst og riktig metabolsk funksjon.
Selv om hormoner, vekstfaktorer og signalmolekyler er avgjørende for riktig proliferasjon av bestemte cellelinjer, bør følgende forholdsregler alltid tas: Siden tilsetning av kosttilskudd kan endre osmolaliteten i det komplette vekstmediet, noe som kan hemme celleutviklingen, anbefales det alltid å kontrollere osmolaliteten etter at kosttilskuddene er tilsatt. For de fleste cellelinjer ligger den optimale osmolaliteten mellom 260 og 320 mOSM/kg.
Antibiotika
Antibiotika brukes ofte for å hemme utviklingen av bakterielle og soppbaserte forurensninger [16], selv om de ikke er avgjørende for cellevekst. Siden antibiotika kan skjule forurensning av mykoplasma og resistente bakterier, anbefales ikke rutinemessig bruk av dem i cellekulturer [17, 18].
I tillegg kan antibiotika forstyrre stoffskiftet i overfølsomme celler. Kombinasjonene av penicillin og streptomycin produsert av MilliporeSigma og Life Technologies brukes ofte. Plasmocin har blitt brukt i dyrking av gliomcellelinjene TS603, TS516 og BT260 [19], og det har vist seg å være effektivt for å fjerne mykoplasmakontaminering (20).
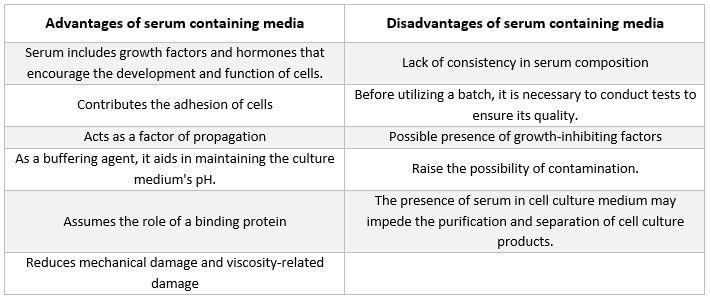
Serum
Albuminer, vekstfaktorer og veksthemmere finnes alle i serum. Serum er en av de viktigste komponentene i cellekulturmedium fordi det tilfører aminosyrer, proteiner, vitaminer (spesielt fettløselige vitaminer som A, D, E og K), karbohydrater, lipider, hormoner, vekstfaktorer, mineraler og sporstoffer.
Serum fra foster og kalv av storfe brukes ofte for å fremme utviklingen av dyrkede celler. Fosterserum er en rik kilde til vekstfaktorer og egner seg for cellekloning og utvikling av følsomme celler. På grunn av dets reduserte evne til å fremme vekst, brukes kalveserum i kontaktinhibisjonsforsøk. Vanlige vekstmedier inneholder ofte 2–10 % serum. Tilsetning av serum til dyrkningsmediet tjener følgende formål [21]:
-
Serumet tilfører cellene essensielle næringsstoffer (både i oppløsning og bundet til proteiner).
-
Serumet inneholder flere vekstfaktorer og hormoner som er involvert i vekstfremmende prosesser og spesialisert celleaktivitet.
-
Det inneholder mange bindingsproteiner, som albumin og transferrin, som transporterer andre kjemikalier inn i cellen. For eksempel transporterer albumin fett, vitaminer, hormoner osv. inn i cellene.
-
Det tilfører også proteiner, som fibronektin, som øker cellenes vedheft til underlaget. I tillegg produserer det spredningselementer som bidrar til celleutvidelse før deling.
-
Det leverer proteasehemmere som forhindrer proteolyse i celler.
-
Det inneholder også mineraler som Na+, K+, Zn2+ og Fe2+.
-
Det øker viskositeten i mediet og beskytter dermed cellene mot mekanisk skade under omrøring i suspensjonskulturen.
-
Det fungerer også som en buffer.
Referanser
[1] Morgan J, Morton H, Parker R. Ernæring av dyreceller i vevskultur; innledende studier av et syntetisk medium. Proc Soc Exp Biol Med. 1950;73:1–8
[2] Kerbel R, Blakeslee D. Rask adsorpsjon av en komponent i føtal kalveserum av pattedyrceller i kultur. En potensiell kilde til artefakter i studier av antiserum mot cellespesifikke antigener. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. Tilsetning av serum til mediet brukt til fremstilling av cellesuspensjoner som en mulig kilde til artefakter i cellemedierte reaksjoner undersøkt ved hjelp av popliteal lymfeknute-testen. J Immunogenet. 1980;7:483-9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Kommersielle serumfrie medier: hybridomvekst og produksjon av monoklonale antistoffer. J Immunol Methods. 1991;145:175-83
[5] Barnes D, Sato G. Metoder for dyrking av celler i serumfritt medium. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O, m.fl. HSP70-chaperoner transporterer RNA-fritt TDP-43 inn i anisotrope, intranukleære, flytende sfæriske skall. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, m.fl. Downsyndrom-indusert senesens forstyrrer kjernestrukturen til nevrale stamceller. Cell Stem Cell. 2022;29:116–130.e7
[8] Iscove N, Melchers F. Fullstendig erstatning av serum med albumin, transferrin og soyalipid i kulturer av lipopolysakkarid-reaktive B-lymfocytter. J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Systematisk forbedring av et kjemisk definert proteinfritt medium for hybridomvekst og produksjon av monoklonale antistoffer. J Biotechnol. 1996;45:111-23
[10] Darfler F. Et proteinfritt medium for vekst av hybridomer og andre celler i immunsystemet. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Serumfri cellekultur: en helhetlig tilnærming. Cell. 1980;22:649-55
[12] Hamilton W, Ham R. Klonal vekst av kinesiske hamstercellelinjer i proteinfritt medium. In Vitro. 1977;13:537-47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Analyse av de cytotoksiske effektene av lysutsatt HEPES-holdig dyrkningsmedium. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. Fenolrødt i vevskulturmedier er et svakt østrogen: implikasjoner for studiet av østrogenresponsive celler i kultur. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Utvikling av serumfrie medier. I: Master JRW, red. Animal Cell culture, 3. utg. Oxford: Oxford University Press; 2000.
[16] Perlman D. Bruk av antibiotika i cellekulturmedier. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. Spredning og bekjempelse av mykoplasmainfeksjon i cellekulturer. In Vitro. 1976;12:643-8
[18] Masters J, Stacey G. Bytte av medium og passering av cellelinjer. Nat Protoc. 2007;2:2276–84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M, et al. Histon-demetylase KDM6A registrerer oksygen direkte for å regulere kromatin og cellenes skjebne. Science. 2019;363:1217-1222
[20] Molla Kazemiha V, Azari S, Amanzadeh A, Bonakdar S, Shojaei Moghadam M, Habibi Anbouhi M, m.fl. Effektiviteten av Plasmocin™ på ulike pattedyrcellelinjer infisert av mollicutes sammenlignet med vanlige antibiotika i cellekultur: en lokal erfaring. Cytotechnology. 2011;63:609-20
[21] Kragh Hansen U. Molekylære aspekter ved ligandbinding til serumalbumin. Pharmacol Rev. 1981;33:17-53