
, L-alaniin-L-glutamiini lahus")
Maht: 100 ml
Säilitamine: ≤ –15 °C
Steriilsus: steriilselt filtreeritud
Stabiilne glutamiinilahus (L-alanüül-L-glutamiin, 200 mM) on väga stabiilne L-glutamiini dipeptiidvorm, mis on loodud asendama tavapärast L-glutamiini rakukultuurikeskkonnas. L-glutamiin on oluline aminohape ja peamine energiaallikas kultiveeritud rakkudele, mängides olulist rolli rakkude kasvu, ainevahetuse ja valgusünteesi protsessides.
Kasutamine ja eelised
Tavalistes vedelates keskkondades laguneb L-glutamiin 37 °C juures suhteliselt kiiresti, mille tulemusena tekivad mürgised kõrvalsaadused, nagu ammooniumioonid, mis võivad negatiivselt mõjutada rakkude eluvõimelisust ja eksperimendi tulemusi. Stabiilne glutamiin ületab selle piirangu, pakkudes lagunematu dipeptiidvormi, mis jääb kultiveerimistingimustes muutumatuks.
Rakud lõhustavad dipeptiidsidet ensümaatiliselt, et vabastada L-glutamiini vastavalt vajadusele, tagades pideva värske varustuse ja vältides samal ajal kahjulike jääkproduktide kogunemist. See muudab lahuse eriti soodsaks pikaajaliste rakukultuuride ja suure tihedusega kasvatussüsteemide jaoks.
Koostis ja kasutus
L-alanüül-L-glutamiini lahendus valmistatakse rakukultuuri jaoks sobivas vees kontsentratsiooniga 200 mM ja see on steriliseeritud filtreeritud saastumisele tundlike rakenduste jaoks. Seda saab lahjendada otse täielikku kasvukeskkonda vastavalt eksperimendi nõuetele. Säilitada temperatuuril ≤ –15 °C ja vältida korduvaid külmutus-sulatus tsükleid, et säilitada toote stabiilsus.
Ainult teadusuuringuteks. Ei ole mõeldud kasutamiseks diagnostilistes või terapeutilistes protseduurides. Ei ole mõeldud kasutamiseks inimestel ega loomadel.
")
Maht: 100 ml
Säilitamine: +2 °C kuni +8 °C
Steriilsus: steriilselt filtreeritud
HEPES-puhverlahus (1 M), tuntud ka kui N-2-hüdroksüetüülpiperasiin-N-2-etaanesulfoonhape, on tsükliline orgaaniline puhveraine, mida kasutatakse laialdaselt rakukultuurikeskkonnas. See on mõeldud pH-taseme stabiilsena hoidmiseks füsioloogilises vahemikus 6,7–8,6, toetades optimaalset rakufunktsiooni in vitro rakendustes.
Kasutamine ja eelised
HEPES tagab usaldusväärse puhvri võime rakukultuuri süsteemides, eriti kui rakke käideldakse väljaspool CO₂ inkubaatorit. 10–25 mM HEPESi lisamine kultuurikeskkonnale tagab parema pH stabiilsuse pikemaajalise käitlemise ajal, aidates säilitada ühtlased katsetingimused.
See puhver on membraanile läbilaskmatu, mõjutab biokeemilisi reaktsioone minimaalselt ja on keemiliselt ja ensümaatiliselt väga stabiilne. Need omadused muudavad selle sobivaks mitmesugusteks rakukultuuri ja biokeemilisteks rakendusteks.
Koostis ja kasutamine
Lahus tarnitakse 1 M kontsentraadina, mis on valmistatud rakukultuuri jaoks sobivas vees ja on steriilselt filtreeritud kasutamiseks saastumisele tundlikes keskkondades. Seda saab lahjendada soovitud töökontsentratsioonini sõltuvalt rakenduse nõuetest. Säilitada +2 °C kuni +8 °C juures ja käsitseda aseptilistes tingimustes, et säilitada toote terviklikkus.
Ainult teadusuuringuteks. Ei ole mõeldud kasutamiseks diagnostilistes või terapeutilistes protseduurides. Ei ole mõeldud kasutamiseks inimestel ega loomadel.
, 250 g/l")
Maht: 50 ml
Säilitamine: +2 °C kuni +8 °C
Steriilsus: steriilselt filtreeritud
D-(+)-glükoosilahus (dekstroosilahus) on steriilne, kasutusvalmis toidulisand, mis sisaldab looduslikult esinevat suhkrut D-(+)-glükoosi, mis on rakkude ainevahetuse keskne komponent. Glükoos osaleb olulistes bioloogilistes protsessides, nagu energia tootmine, glükosüülimine ja glükaanide moodustumine, mis aitavad kaasa rakkude struktuuri ja funktsioneerimisele.
Kasutamine ja eelised
Seda glükoosilahust kasutatakse laialdaselt lisandina rakukultuurikeskkonnas ja paljudes raku
- ja molekulaarbioloogia rakendustes. Glükoos on peamine süsiniku
- ja energiaallikas, mis toetab rakkude kasvu, paljunemist ja ainevahetust. Tema osalemine biosünteetilistes protsessides muudab ta oluliseks normaalse rakufüsioloogia ja eksperimentaalse järjepidevuse säilitamisel.
Koostis ja kasutamine
Lahus on saadaval kõrge kontsentratsiooniga 250 g/l glükoosi, mis võimaldab seda kultuurikeskkonnas eksperimendi nõuetele vastavalt paindlikult lahjendada. See on steriilselt filtreeritud, et tagada sobivus saastumisele tundlikeks rakendusteks. Säilitada +2 °C kuni +8 °C juures ja käsitseda aseptiliselt, et säilitada toote kvaliteet ja stabiilsus.
Ainult teadusuuringuteks. Ei ole mõeldud kasutamiseks diagnostilistes või terapeutilistes protseduurides. Ei ole mõeldud kasutamiseks inimestel ega loomadel.
, lahus 100x")
Maht: 10 ml
Säilitamine: +2 °C kuni +8 °C
Sterilsus: steriilselt filtreeritud
Insuliin-transferriin-seleen (ITS) lahus (100x) on keemiliselt määratletud täiendusaine, mis on mõeldud laiale valikule rakukultuuri rakendustele. Seda kasutatakse kõige sagedamini lisandina rakukultuuri baaskeskkonnale, et toetada rakkude kasvu vähendatud seerumi või seerumivabas keskkonnas.
Kasutamine ja eelised
Meie ITS-lisand pakub olulisi komponente, mis on vajalikud seerumivaba keskkonna optimaalseks toimimiseks. Täiendades tavapärast toitainekeskkonda ITS-iga, saab oluliselt vähendada loote veise seerumi (FBS) vajadust paljude rakuliinide rutiinseks hooldamiseks. See aitab vähendada seerumi kasutamisega seotud variatiivsust, säilitades samal ajal rakkude kasvu ja eluvõime.
Insuliin toetab oluliste toitainete rakkudesse imendumist ja ainevahetust, transferriin hõlbustab raua transporti ning seleen aitab kaasa antioksüdantkaitsele ja ensüümiaktiivsusele. Koos soodustavad need komponendid tasakaalustatud rakuainevahetust ja paremat reprodutseeritavust määratletud kultuursüsteemides.
Koostis ja kasutamine
Insuliin-transferriin-seleen (ITS) tarnitakse 100-kordselt kontsentreeritud lahendusena Earle'i tasakaalustatud soolalahuses (EBSS) ilma fenoolpunaseta. Standardrakenduste puhul lahjendage 1:100 sobivasse baaskeskkonnas, et saavutada soovitatav töökontsentratsioon. Hoida temperatuuril +2 °C kuni +8 °C ja käsitseda aseptilistes tingimustes, et säilitada toote stabiilsus ja steriilsus.
Ainult teadusuuringuteks. Ei tohi kasutada diagnostilistes ega terapeutilistes protseduurides. Ei tohi kasutada inimestel ega loomadel.
")
Maht: 5 ml
Säilitamine: +2 °C kuni +8 °C
Sterilsus: steriilselt filtreeritud
Inimese rekombinantse insuliini lahus on keemiliselt määratletud täiendusaine, mida kasutatakse tavaliselt imetajate rakuliinide, sealhulgas hiina hamstri munasarja (CHO) rakkude kasvatamiseks. See rakukultuuri jaoks mõeldud lahus sisaldab Saccharomyces cerevisiae's ekspresseeritud rekombinantse inimese insuliini, mis tagab kõrge puhtuse ja stabiilse toimivuse teadusuuringutes.
Kasutamine ja eelised
Insuliini lisatakse tavaliselt seerumivabadele ja keemiliselt määratletud kasvukeskkondadele, et soodustada rakkude kasvu ja produktiivsust. Insuliin on oluline regulatiivne hormoon, mis toetab glükoosi, aminohapete ja rasvhapete rakkudesse imendumist, kasutamist ja ladustamist. Samuti pärsib see glükogeeni, valkude ja lipiidide lagunemist, aidates seeläbi parandada rakkude eluvõimelisust ja ainevahetuse stabiilsust kultuurisüsteemides. Keemiliselt määratletud koostis toetab reprodutseeritavust ja vähendab tundlike rakukultuuri töövoogude varieeruvust.
Bioloogilised omadused ja kasutus
Insuliin on kahe ahelaga polüpeptiidhormoon, mida toodavad loomulikult pankrease saarekeste β-rakud. Selle molekulmass on umbes 5800 Da. α
- ja β-ahelad on ühendatud kahe ahelatevahelise disulfiidsidemega ning α-ahel sisaldab ühte ahelasisest disulfiidsidet. Rakukultuuri rakenduste puhul tuleb lahust käsitseda aseptilistes tingimustes ja säilitada temperatuuril +2 °C kuni +8 °C, et säilitada stabiilsus ja toimivus.
Ainult teaduslikuks kasutamiseks. Ei tohi kasutada diagnostilistes ega terapeutilistes protseduurides. Ei tohi kasutada inimestel ega loomadel.
")
Maht: 100 ml
Säilitamine: +2 °C kuni +8 °C
Steriilsus: steriilselt filtreeritud
Naatriumpüruvaadi lahus (100 mM) on steriilne, kasutusvalmis lisand, mis on mõeldud rakkude kasvatuskeskkondadele täiendava, kergesti kättesaadava energiaallika pakkumiseks. Naatriumpüruvaat mängib olulist rolli rakkude energiaainevahetuses ja toetab metaboolselt aktiivsete ja kiiresti paljunevate rakkude, näiteks kasvajarakkude kasvu. Lisand võib parandada rakkude eluvõimelisust ja aidata säilitada metaboolset stabiilsust kasvatussüsteemides.
Kasutamine ja eelised
Seda lahust kasutatakse laialdaselt rutiinsetes rakukultuurides, et rikastada keskkonda püruvaadiga ja soodustada optimaalseid kasvutingimusi. See toetab ATP tootmist, võib aidata vähendada oksüdatiivset stressi ja aitab parandada kultiveeritud rakkude ainevahetust. Toode on valmistatud rakukultuurikvaliteediga veest ja steriilselt filtreeritud, tagades uurimistööde käigus ühtlase kvaliteedi ja korratavuse.
Kasutamine ja ühilduvus
Enamiku rakukultuuri rakenduste soovitatav lõplik kontsentratsioon on 1 mM naatrium-püruvaat, mis saavutatakse 100 mM lahusega lahjendades 1:100 täielikku kultuurikeskkonna. Lahus on ühilduv paljude baaskeskkonna ja imetajate rakuliinidega. Säilitage +2 °C kuni +8 °C juures ja kaitse saastumise eest, et säilitada toote stabiilsus.
Ainult teadusuuringuteks. Ei ole mõeldud kasutamiseks diagnostilistes või terapeutilistes protseduurides. Ei ole mõeldud kasutamiseks inimestel ega loomadel.
")
Maht: 100 ml Säilitamine: ≤-15°C Steriilsus: Steriilselt filtreeritud
Antibiootikum/antimükootiline lahus (100x) on steriilne, kasutusvalmis kontsentraat, mis on mõeldud mikroobse saastumise riski vähendamiseks rakukultuurides ja nendega seotud laboratoorsetes rakendustes. See 100x lahus sisaldab penitsilliini, streptomütsiini ja amfoteritsiin B hästi tõestatud kombinatsiooni, mis tagab laia spektriga antimikroobse toime grampositiivsete ja gramnegatiivsete bakterite, pärmseente ja filamentsete seente vastu. Preparaat sobib kasutamiseks eukarüootilistes rakukultuurides, bakteriaalsetes keskkondades ja muudes saastetundlikes süsteemides, toetades puhtaid ja järjepidevaid laboritoiminguid.
Kasutamine ja eelised Seda rutiinsete teadusprotokollide jaoks optimeeritud lahust kasutatakse laialdaselt aseptiliste tingimuste säilitamiseks rakukultuuride tööprotsessides. See pakub usaldusväärset toimimist saastetundlikes keskkondades, aidates teadlastel vähendada mikroobide ülekasvu ohtu, ilma et see kahjustaks rakkude tervist või katsete reprodutseeritavust. Steriilne filtreeritud koostis välistab vajaduse täiendavate solubiliseerimisetappide järele, toetades ühtlustatud söötme ettevalmistamist ja vähendades igapäevaste laboriprotseduuride varieeruvust.
Kasutamine ja ühilduvus Standardsete töökontsentratsioonide saavutamiseks lahjendage lahust 1:100 teie täielikule kultuurikeskkonnale. Toode ühildub paljude imetajate rakuliinide ja põhikeskkondadega. Tänu pidevale varude kättesaadavusele saavad teadlased kasu usaldusväärsest tarnekindlusest ja lihtsustatud logistika planeerimisest. Stabiilsuse säilitamiseks tuleb lahust hoida temperatuuril ≤ -15 °C ja kaitsta korduvate külmutamis
- ja sulatamistsüklite eest. Ainult teaduslikuks kasutamiseks. Mitte kasutamiseks diagnostilistes või terapeutilistes protseduurides. Mitte kasutamiseks inimestel või loomadel.
")
Maht: 100 ml Säilitamine: +2°C kuni +8°C Steriilsus: Steriilne filtreeritud
MEM Non-Essential Amino Acids (100x) on steriilne toidulisand, mis on mõeldud rakkude kasvu ja elujõulisuse suurendamiseks imetajate rakukultuurisüsteemides. Koostis vastab 100x kontsentraadile mitteolemuslikest aminohapetest, mida leidub standardses minimaalses asendamatus keskkonnas (MEM), võimaldades baasmeediumide otsest täiendamist minimaalse ettevalmistusega.
Kasutamine ja eelised See täiendus annab täiendava aminohapete reservi kiiresti proliferatsiooni alustavatele rakkudele või rakuliinidele, mis on kaotanud võime sünteesida mitteolulisi aminohappeid de novo. Vähendades biosünteesi metaboolset koormust, toetab see paremat kasvukineetikat, pikemat elujõulisust ja suuremat katsete järjepidevust
- eriti toitainete suhtes tundlikes või suure tihedusega kultuurides.
Koostis ja kasutamine Lahus sisaldab glütsiini, L-alaniini, L-asparagiini, L-asparagiinhapet, L-glutamiinhapet, L-proliini ja L-seriini. See sobib kokku MEM-i ja enamiku teiste standardkeskkondadega. Kasutamiseks lahjendatakse 1:100 lõplikus kultuurikeskkonnas. See toode on steriilselt filtreeritud ja kasutusvalmis ilma täiendavate käitlemisetappideta. Ainult teaduslikuks kasutamiseks. Mitte kasutamiseks diagnostilistes või terapeutilistes protseduurides. Mitte kasutamiseks inimestel või loomadel.

Accutase on kasutusvalmis, steriilselt filtreeritud rakkude eralduslahus, mis on loodud kui õrn alternatiiv trüpsiinile/EDTA-le imetajate adhesiivsete rakkude eraldamiseks standardsetest kudekultuuri plastnõudest ja adhesiivkattega pindadelt. See ühendab proteolüütilise ja kollageenolüütilise ensüümi aktiivsuse tasakaalustatud soolalahuses, et tagada efektiivne, kuid kontrollitud eraldamine, säilitades rakupinna valgud ning toetades kõrget eluvõimet pärast passaaži ja kiiret uuesti kinnitumist.
Accutase koostis põhineb Dulbecco fosfaadipuhverdatud soolalahusel (DPBS), millele on lisatud EDTA ja fenoolpunane visuaalse pH indikaatorina. Ensüümid on mitte-imetajate ja mitte-bakteriaalset päritolu, mistõttu Accutase sobib eriti hästi tüvirakkude uurimiseks, vaktsiinide tootmiseks ja mis tahes rakendusteks, kus tuleb minimeerida loomsetest või mikroobidest pärinevaid saasteaineid. Lahus inhibeerib ennast automaatselt 37 °C juures, seega ei ole pärast eraldamist vaja neutraliseerivat reagenti ega seerumit sisaldavat keskkonda – rakud saab viia otse värskesse keskkonda.
Peamised omadused
Kasutusvalmis 1x steriilselt filtreeritud vedelik – lahjendamist ega taastamist ei ole vaja
Kombineeritud proteolüütiline ja kollageenolüütiline ensüümiaktiivsus õrnaks dissotsiatsiooniks
Iga partii on standardiseeritud kindlaksmääratud dissotsiatsioonitegevusele, et tagada partiidevaheline ühtlus
Enüümid ei pärine imetajatelt ega bakteritelt
Inhibeerib ennast 37 °C juures – neutraliseerivat lahust ei ole vaja
Valmistatud Dulbecco PBS-is koos EDTA-ga
Fenoolpunane on lisatud visuaalseks pH-indikaatoriks
pH 6,8–7,8
Tüüpilised rakendused
Accutase lahustab õrnalt mitmesuguseid adhesiivseid ja tundlikke rakutüüpe, sealhulgas inimese embrüonaalseid tüvirakke (hESCs), inimese indutseeritud pluripotentseid tüvirakke (iPSCs), närvi tüvirakke, primaarseid neuroneid ja tavapäraselt kultiveeritavaid adhesiivseid rakuliine, nagu HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 ja A549. Tüüpilised kasutusjuhtumid hõlmavad:
Imetajate adhesiivsete rakkude rutiinne subkultiveerimine ja passaažimine
hESCs, iPSCs ja muude tundlike liinide õrn üherakuline dissotsiatsioon
Proovide ettevalmistamine voolutsütomeetria ja FACS-analüüsi jaoks
Rakupinna markerite analüüs, kus epitoobi terviklikkus on oluline
Rakkude rändamise, proliferatsiooni ja apoptoosi analüüsid
Puhkeoleku testid seerumi näljutamise ja onkogeeni transfektsiooni uuringute abil
Tuumorrakkude ja neuraalkreestrakkude rändetestid
Tootmise mastaabis laiendamine bioreaktori töövoogudes
Rutiinseks tööks kandke umbes 10 ml Accutase'i 75 cm2 kultuuripinnale ja inkubeerige 5–10 minutit toatemperatuuril. Optimaalne inkubatsiooniaeg tuleks määrata iga rakuliini jaoks eraldi ja see ei tohiks ületada ühte tundi. Enne lisamist loputage rakukihiga Ca2+/Mg2+-vaba soolalahusega, näiteks kaltsiumi ja magneesiumita DPBS-ga, et eemaldada jääkserum ja kahevalentsed katioonid.
Käsitsemine ja säilitamine
Hoidke avamata pudelit külmutatuna temperatuuril -15 °C või madalamal. Sulatage toatemperatuuril või öösel temperatuuril +2 °C kuni +8 °C. Ärge sulatage Accutase'i 37 °C veevannis, kuna kõrgem temperatuur vähendab ensüümi aktiivsust. Pärast sulatamist võib lahust hoida kuni 2 kuud temperatuuril +2 °C kuni +8 °C; ärge hoidke toatemperatuuril. Ärge soojendage reagenti enne kasutamist 37 °C-ni – lisage see otse pestud rakkudele toatemperatuuril. Pika säilivusaja tagamiseks soovitatakse kasutada ühekordseks kasutamiseks mõeldud alikvoote, et vältida korduvaid sulatamistsükleid. Töötage alati aseptilistes tingimustes.
Kvaliteet
Valmistatud rangete kvaliteedistandardite kohaselt. Iga Accutase'i partii on steriilselt filtreeritud ja testitud steriilsuse, pH, välimuse ja dissotsiatsioonitegevuse suhtes, et tagada partii-partii ühtlane ja korratav tulemus.
Toote spetsifikatsioonid
Spetsifikatsioon
Detail
TootetüüpRakkude eraldamise / dissotsiatsiooni reagent
VormSteriilselt filtreeritud vedelik, kasutusvalmis
Maht100 ml
Töökontsentratsioon1x (kasutusvalmis)
Enüümi aktiivsusKombineeritud proteolüütiline ja kollageenolüütiline
Enüümi päritoluMitte-imetaja ja mitte-bakteriaalne
PuhversüsteemDulbecco PBS koos EDTA-ga
pH indikaatorFenoolpunane
pH vahemik6,8 – 7,8
VälimusLäbipaistev, heleroosa kuni oranž lahus
Säilitustemperatuur-15 °C või madalam
Stabiilsus pärast sulatamistKuni 2 kuud temperatuuril +2 °C kuni +8 °C
Soovitatav kasutamiskogus~10 ml 75 cm2 kultuuripinna kohta
Tüüpiline inkubatsiooniaeg5–10 minutit toatemperatuuril
TransporditingimusedKülmutatud kuivjääs
KasutuseesmärkAinult teadusuuringuteks ja edasiseks tootmiseks
Koostis (koostis ühe liitri kohta)
Komponent
Kontsentratsioon (mg/l)
Anorgaanilised soolad
Naatriumkloriid (NaCl)8000,00
Dinatriumvesinikfosfaat (Na2HPO4)1150,00
Kaaliumkloriid (KCl)200,00
Kaaliumdivesinikfosfaat (KH2PO4)200,00
Muud koostisosad
EDTA · 4Na (tetranatrium-EDTA)220,00
Fenoolpunane3,00
Patenteeritud ensüümisegu (proteolüütiline ja kollageenolüütiline toime)1x
Accutase on Innovative Cell Technologies, Inc. registreeritud kaubamärk.
, w: 2 mM L-Glutamiin, w: 2,2 g/L NaHCO3, w: EBSS")
Seda kasutusvalmis, steriilselt filtreeritud vedelat koostist on täiendatud Earle’i tasakaalustatud soolalahusega (EBSS), 2 mM L-glutamiiniga, D-glükoosiga (1,0 g/l) ja 2,2 g/l naatriumvesinikkarbonaadiga (NaHCO3), mis muudab selle sobivaks kasutamiseks CO2-kontrollitud inkubaatori keskkonnas (tavaliselt 5 % CO2). Lisatud fenoolpunane toimib pH-indikaatorina, võimaldades rakukultuuri ajal keskkonna seisundi mugavat visuaalset jälgimist.
Peamised omadused
Klassikaline Eagle’i MEM-koostis Earle’i tasakaalustatud soolalahusega (EBSS)
Sisaldab 2 mM L-glutamiini – valmis koheseks kasutamiseks
2,2 g/l naatriumvesinikkarbonaati – puhverdatud 5 % CO2 inkubeerimiseks
Peamise süsinikuallikana D-glükoos (1,0 g/l)
Fenoolpunane pH-indikaatorina
Ilma HEPES-ita ja ilma naatrium-püruvaadita
Steriilselt filtreeritud vedel keskkond, valmis kasutamiseks
pH 7,0 – 7,6
Tüüpilised rakendused
EMEM toetab mitmesuguste imetajate rakuliinide kasvatamist, sealhulgas HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 ja paljusid esmaseid rakke. Tüüpilised rakendused hõlmavad:
Adherentse rakuliini rutiinne hooldus ja paljundamine
Viiruste paljundamine ja vaktsiinide tootmine
Tsütotoksilisuse ja biotestide rakendused
Transfektsiooni ja valkude ekspressiooni uuringud
Alusuuringud rakubioloogias ja molekulaarbioloogias
Optimaalse rakkude kasvu tagamiseks lisatakse EMEM-ile tavaliselt 5–10 % loote veise seerumit (FBS) ja, sõltuvalt rakuliinist, mitte-essentsiaalseid aminohappeid (NEAA) ning antibiootikume, nagu penitsilliin/streptomütsiin.
Käsitsemine ja ladustamine
Hoida avamata pudelit +2 °C kuni +8 °C juures, valguse eest kaitstult. Pärast avamist kasutada aseptilistes tingimustes. Lahuses olev L-glutamiin laguneb järk-järgult – parima tulemuse saavutamiseks soovitame kasutada keskkonda 4 nädala jooksul pärast avamist või lisada enne kasutamist värsket L-glutamiini, kui seda hoitakse kauem. Enne rakkudele lisamist lasta keskkonnal soojeneda 37 °C-ni.
Kvaliteet
Valmistatud rangete kvaliteedistandardite kohaselt. Iga partii steriilsust, pH-d, osmolalisust ja endotoksiinitaset kontrollitakse, et tagada ühtlane toimivus rakukultuuri rakendustes.
Toote spetsifikatsioonid
Spetsifikatsioon
Detail
Toote tüüpMEM
TootekategooriaRakukultuuri kasvukeskkonnad
VormVedelik
SteriilneJah
Suurus500 ml
L-glutamiinL-glutamiiniga (2 mM)
GlükoosGlükoosiga (1,0 g/l)
NaHCO3NaHCO3-ga (2,2 g/l)
HEPESIlma HEPES-ita
NaHCO3Ilma naatrium-püruvaadita
FenoolpunaneFenoolpunasega
SoolalahusEarle’i tasakaalustatud soolalahus (EBSS)
pH7,0 – 7,6
EndotoksiinisisaldusEi ole täpsustatud
Säilitamine+2 °C kuni +8 °C
Koostis (koostis ühe liitri kohta)
Komponent
Kontsentratsioon (mg/l)
Anorgaanilised soolad
Kaltsiumkloriid · 2H2O265,00
Magneesiumsulfaat97,72
Kaaliumkloriid400,00
Naatriumkloriid6 800,00
Na-divesinikfosfaat, veevaba122,00
Naatriumvesinikkarbonaat (NaHCO3)2 200,00
Aminohapped
L-arginiin · HCl126,00
L-tsüstiin · 2HCl31,30
L-glutamiin292,00
L-histidiin · HCl · H2O42,00
L-isoleutsiin52,00
L-leutsiin52,00
L-lüsiin · HCl72,50
L-metioniin15,00
L-fenüülalaniin32,00
L-treoniin48,00
L-trüptofaan10,00
L-türosiin · 2Na · 2H2O51,90
L-valiin46,00
Vitamiinid
D-kaltsiumpantotenaat1,00
Koliinkloriid1,00
Foolhape1,00
Müo-inositool2,00
Nikotiinamiid1,00
Püridoksaal · HCl1,00
Riboflaviin0,10
Tiamiin · HCl1,00
Muud koostisosad
D(+)-glükoos1 000,00
Fenoolpunane10,00

- kui ka loomsetele rakkudele, muutes selle oluliseks vahendiks mitmesuguste teadusrakenduste jaoks. Freeze Medium CM-1, mis on koostatud hoolikalt tasakaalustatud krüoprotektiivide ja oluliste toitainete kombinatsiooniga, minimeerib jääkristallide moodustumist ja rakkude stressi külmutamise ajal, säilitades seega rakkude terviklikkuse.
Freeze Medium CM-1 põhiomadused on järgmised:
Laiaulatuslik kokkusobivus: Tõhus paljude rakutüüpide, sealhulgas primaarsete rakkude, tüvirakkude ja väljakujunenud rakuliinide puhul.
Kõrge elujõulisus: Optimeeritud selleks, et maksimeerida rakkude taastumist ja elujõulisust pärast sulatamist, tagades usaldusväärsed katsetulemused.
Kasutusvalmis: Mugav valmistada ja steriliseerida koheseks kasutamiseks, vähendades ettevalmistusaega ja saastumisohtu.
Suurendatud stabiilsus: Säilitab järjepideva tulemuslikkuse standardsetes krüokonserveerimistingimustes, tagades reprodutseeritavad tulemused.
Pikk säilivusaeg: CM-1 on seerumit sisaldav, kasutusvalmis krüokonserveerimissöötme, mida võib säilitada külmkapis kuni üks aasta.
CM-1 kasutamine rakkude külmutamiseks
CM-1 kasutamiseks nii adherentsete kui ka suspensioonirakkude külmutamiseks järgige järgmisi samme
Adherentsete rakkude puhul peske ja eraldage need kultuurisubstraadist. Suspensioonirakkude puhul jätkake otse järgmise sammuga.
Arvutage rakke, et tagada nende õige kontsentratsioon.
Pelleteerimiseks tsentrifuugige rakud, seejärel resuspenseerige need CM-1 külmutuskeskkonnas.
Ülekandke resuspendeeritud rakud krüoklaasidesse.
Kasutage enne rakkude pikaajalisse säilitamisse viimist aeglast külmutamist
Meetod
Kirjeldus
Sammud
❄️
Käsitsi külmutamine
Järkjärguline meetod, mis hõlmab järk-järgulist temperatuuri vähendamist rakkude elujõulisuse tagamiseks
1️⃣ Asetage rakud külmutuskeskkonda 4 °C sügavkülmikusse 40 minutiks.
2️⃣ Viiakse 24 tunniks -80 °C sügavkülmikusse.
3️⃣ Säilitada rakke pikaajalise säilitamise eesmärgil vedelas lämmastikus
❄️
Mr. Frosty kasutamine
Mugav seade, mis võimaldab kontrollitud külmutamiskiirust ilma elektrienergiata
1️⃣ Valmistage rakud krüoviaalides külmutuskeskkonnaga ette.
2️⃣ Asetage krüoviaalid Mr. Frosty konteinerisse.
3️⃣ Säilitage 24 tundi temperatuuril -80 °C enne vedela lämmastiku sisse viimist
❄️
Reguleeritava kiirusega sügavkülmik
Thermo Fisheri või teiste tootjate kõrge täpsusega sügavkülmik, mis on ette nähtud temperatuuri kontrollitud langetamiseks
1️⃣ Programmeerige seade nii, et temperatuur langeb järk-järgult.
2️⃣ Asetage ettevalmistatud rakud sügavkülmikusse.
3️⃣ Pärast külmutamistsüklit viige rakud üle vedela lämmastikuga
Säilitage krüoviaalid temperatuuril alla -130 °C või pikaajalise säilitamise eesmärgil vedelas lämmastikus.
Koostisosad
Sisaldab FBS, DMSO, glükoosi, soolasid
Puhverdusvõime: pH = 7,2 kuni 7,6
Cytion's Freeze Medium CM-1 pakub usaldusväärset lahendust krüokonserveerimiseks, tagades rakkude kõrge elujõulisuse ja funktsionaalsuse pärast sulatamist mitmesuguste teadusrakenduste jaoks.

- ja kanamaksa rakkude kasvatamist, eriti vähendatud seerumi tingimustes.
Ham's F-12K (Kaighn's) Medium on hoolikalt koostatud rakukultuuritingimuste optimeerimiseks. Sellel on rikastatud koostis, mis pakub kõrgendatud koguseid olulisi komponente, nagu aminohappeid ja naatriumpüruvaati, samuti täiendavaid elemente, sealhulgas putressiini, tümidiini, hüpoksantiini ja tsinki. Need lisandid võimaldavad teadlastel täiendada keskkonda minimaalse seerumi või konkreetsete rakutüüpide jaoks määratletud komponentidega, hõlbustades täpseid katsetingimusi.
Eelkõige ei sisalda Ham's F-12K (Kaighn's) keskkond valke ega kasvufaktoreid. Sellest tulenevalt on sageli vaja lisada kasvufaktoreid ja veiste loote seerumit (FBS), mis võimaldab teadlastel kohandada keskkonda vastavalt oma konkreetsete rakuliinide nõuetele. Optimaalse tulemuslikkuse tagamiseks tuleb FBS-i kontsentratsioon iga rakuliini jaoks hoolikalt optimeerida, tagades optimaalse kasvu ja funktsionaalsuse.
Füsioloogilise pH säilitamiseks kasutatakse Ham's F-12K (Kaighn's) keskkonnas naatriumbikarbonaatpuhvrisüsteemi (2,5 g/l), mis eeldab kasvatamise ajal kontrollitud 5-10% CO2-keskkonda. See tagab, et keskkonna pH jääb rakkude kasvu ja elujõulisuse jaoks ideaalsesse vahemikku
Kvaliteedikontroll
pH = 7,2 +/
- 0,02 temperatuuril 20-25 °C.
Iga partii on testitud steriilsuse ning mükoplasma ja bakterite puudumise suhtes.
Hooldus
Hoida külmikus +2°C kuni +8°C juures pimedas. Külmutamine ja soojendamine kuni +37°C vähendavad toote kvaliteeti.
Ärge kuumutage keskkonda üle 37°C ega kasutage kontrollimatuid soojusallikaid (nt mikrolaineahjusid).
Kui on vaja kasutada ainult osa söötmest, võtke see kogus pudelist välja ja soojendage seda toatemperatuuril.
Kõikide söötmete, välja arvatud põhisöötme säilivusaeg on 8 nädalat alates tootmiskuupäevast.
Koostis
Koostisosad
mg/l
Anorgaanilised soolad
Kaltsiumkloriid x 2H2O
135,24
Vask(II)-sulfaat x 5H2O
0,00
Raud(II)-sulfaat x 7H2O
0,83
Magneesiumkloriid x 6H2O
105,72
Magneesiumsulfaat x 7H2O
394,49
Kaaliumkloriid
283,29
Kaaliumdivesinikfosfaat
58,52
Naatriumkloriid
7597,20
dinaatriumvesinikfosfaatveevaba
115,02
Tsinksulfaat x 7H2O
0,14
Muud komponendid
D(+)-glükoos veevaba
1260,00
Hüpoksantiin
4,08
DL-α-lipoehape
0,21
Fenool punane
3,00
Putressiin x 2HCl
0,32
Naatriumpüruvaat
220,00
NaHCO3
2500,00
Tümidiin
0,73
Aminohapped
L-alaniin
17,82
L-arginiin x HCl
421,40
L-asparagiin x H2O
30,02
L-asparagiinhape
26,62
L-tsüsteiin x HCl x H2O
70,24
L-glutamiin
292,20
L-glutamiinhape
29,42
Glütsiin
15,01
L-Histidiin x HCl x H2O
41,92
L-isoleutsiin
7,87
L-leutsiin
26,24
L-Lüsiin x HCl
73,04
L-metioniin
8,95
L-fenüülalaniin
9,91
L-proliin
69,06
L-Seriin
21,02
L-treoniin
23,82
L-trüptofaan
4,08
L-Türosiin
10,87
L-valiin
23,42
Vitamiinid
D(+)-Biotiin
0,07
D-kaltsium-pantotenaat
0,48
Koliinkloriid
13,96
Foolhape
1,32
müo-Inositool
18,02
Nikotiinamiid
0,04
Püridoksiin x HCl
0,06
Riboflaviin
0,04
Tiamiin x HCl
0,34
B12-vitamiin
1,36

Fosfaadiga puhverdatud soolalahus (PBS) on bioloogilistes ja keemilistes uuringutes laialdaselt kasutatav puhverlahus. See mängib olulist rolli pH-tasakaalu ja osmolaarsuse säilitamisel erinevate katsete, sealhulgas kudede töötlemise ja rakukultuuride käigus. Meie PBS-lahus on hoolikalt formuleeritud kõrge puhtusastmega koostisosadest, et tagada stabiilsus ja usaldusväärsus igas eksperimendis. Meie PBS-i osmolaarsus ja ioonide kontsentratsioonid jäljendavad täpselt inimkeha omast, mistõttu on see isotooniline ja enamikule rakkudele mittetoksiline.
Meie PBS-lahuse koostis
Meie PBS-lahus on ülipuhaste fosfaatpuhvrite ja soolalahuste pH-ga kohandatud segu. 1X töökontsentratsioonis sisaldab see järgmist:
8000 mg/l naatriumkloriidi (NaCl)
200 mg/l kaaliumkloriidi (KCl)
1150 mg/L naatriumfosfaat kahealuseline veevaba (Na2HPO4)
200 mg/L Kaaliumfosfaat monobaasiline veevaba (KH2PO4)
See koostis tagab optimaalse pH ja ioonitasakaalu, mis sobib mitmesuguste bioloogiliste rakenduste jaoks.
Meie PBS-lahuse rakendused
Meie PBS-lahus sobib ideaalselt mitmesugusteks rakendusteks bioloogilistes uuringutes. Selle isotoonilised ja mittetoksilised omadused muudavad selle sobivaks ainete lahjendamiseks ja rakukonteinerite loputamiseks. EDTA-d sisaldavad PBS-lahused on tõhusad kinnitunud ja klompunud rakkude eraldamiseks. PBS-i ei tohiks siiski lisada kahevalentseid metalle, nagu tsink, kuna see võib põhjustada sadestumist. Sellistel juhtudel soovitatakse kasutada Good's puhvreid. Lisaks on meie PBS-lahus vastuvõetav alternatiiv viiruste transpordikeskkonnale RNA-viiruste, sealhulgas SARS-CoV-2 transportimiseks ja säilitamiseks.
Kvaliteedikontroll
Steriilselt filtreeritud
Säilitamine ja säilivusaeg
Säilitada +2°C kuni +25°C juures, valguse eest kaitstult.
Pärast avamist säilitada temperatuuril 2°C kuni 25°C ja kasutada 24 kuu jooksul.
Veotingimused
Keskkonnatemperatuur
Hooldus
Hoida külmkapis +2°C kuni +8°C juures pimedas. Vältida külmutamist ja sagedast soojendamist kuni +37°C, kuna see vähendab toote kvaliteeti.
Ärge kuumutage keskkonda üle 37°C ega kasutage kontrollimatuid soojusallikaid, näiteks mikrolaineahjusid.
Kui kasutatakse ainult osa söötmest, võtke vajalik kogus välja ja soojendage see enne kasutamist toatemperatuurini.
Koostis
Kategooria
Komponendid
Kontsentratsioon (mg/l)
Soolad
Kaaliumkloriid
200
Kaaliumfosfaat monobaasiline veevaba
200
Naatriumkloriid
8000
Naatriumfosfaat kahealuseline veevaba
1150

Esialgu inimleukeemiarakkude kasvu toetamiseks nii suspensiooni
- kui ka monokihiliste kultuuride puhul loodud RPMI 1640 keskkond on teadlaste ja kaubanduslike tarnijate muudatuste kaudu muutunud sobivaks mitmesuguste imetajarakkude jaoks. See sobib erakordselt hästi selliste rakuliinidega nagu HeLa, Jurkat, MCF-7, PC12, PBMC, astrotsüüdid ja kartsinoomid.
RPMI 1640 Medium eristub teistest rakukultuurikeskkondadest oma unikaalse koostise poolest. See sisaldab märkimisväärses koguses fosfaate, aminohappeid ja vitamiine. Eelkõige sisaldab see biotiini, B12-vitamiini ja PABA-d, mis puuduvad Eagle's Minimal Essential Medium'is või Dulbecco's Modified Eagle Medium'is. Lisaks sellele on RPMI 1640 Mediumis oluliselt kõrgemad inositooli ja koliini vitamiinide kontsentratsioonid. See ei sisalda aga valke, lipiide ega kasvufaktoreid. Sellest tulenevalt on rakkude optimaalsete kasvutingimuste tagamiseks tavaliselt vaja lisada 10% veiste loote seerumit (FBS).
RPMI 1640 keskkonda puhverdav süsteem põhineb naatriumbikarbonaadil ja vajab 5-10% CO2 keskkonda, et säilitada füsioloogiliselt sobiv pH. Redutseeriva aine glutatiooni lisamine eristab seda keskkonda teistest veelgi.
Kvaliteedikontroll
Steriilselt filtreeritud
Säilitamine ja säilivusaeg
Säilitada +2°C kuni +8°C juures, valguse eest kaitstult.
Pärast avamist säilitada 4°C ja kasutada 6-8 nädala jooksul.
Veotingimused
Keskkonnatemperatuur
Hooldus
Hoida külmkapis +2°C kuni +8°C juures pimedas. Vältida külmutamist ja sagedast soojendamist kuni +37°C, kuna see vähendab toote kvaliteeti.
Ärge kuumutage keskkonda üle 37°C ega kasutage kontrollimatuid soojusallikaid, näiteks mikrolaineahjusid.
Kui kasutatakse ainult osa söötmest, võtke vajalik kogus välja ja soojendage see enne kasutamist toatemperatuurini.
Koostis
Kategooria
Komponendid
Kontsentratsioon (mg/l)
Aminohapped
Glütsiin
10.00
L-Alanüül-L-glutamiin
434.40
L-arginiin
200.00
L-asparagiinH2O
56.82
L-asparagiinhape
20.00
L-tsüstiin 2HCl
65.20
L-glutamiinhape
20.00
L-Histidiin HClH2O
20.27
L-Hüdroksü-L-proliin
20.00
L-isoleutsiin
50.00
L-leutsiin
50.00
L-Lüsiin HCl
40.00
L-metioniin
15.00
L-Fenüülalaniin
15.00
L-proliin
20.00
L-Seriin
30.00
L-treoniin
20.00
L-trüptofaan
5.00
L-Türosiin 2Na 2H2O
28.83
L-Valiin
20.00
Vitamiinid
p-Aminobensoehape
1.00
D-Biotiin
0.20
Koliinkloriid
3.00
D-kaltsium-pantotenaat
0.25
Foolhape
1.00
myo-Inositool
35.00
Nikotiinamiid
1.00
Püridoksiin HCl
1.00
Riboflaviin
0.20
Tiamiini HCl
1.00
B12-vitamiin
0.005
Anorgaanilised soolad
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Muud komponendid
D-glükoos
2000.00
L-glutatioon redutseeritud
1.00
Fenoolpunane naatriumsool
5.30
, w: 3,1 g/L glükoosi, w: 2,5 mM L-glutamiini, w: 15 mM HEPES, w: 0,5 mM naatriumpüruvaati, w: 1,2 g/L NaHCO3")
See unikaalne koostis ühendab Dulbecco modifitseeritud Eagle'i keskkonna (DMEM) ja Ham's F-12 (Ham's Nutrient Mixture F-12) täpses 1:1 suhtes. L-glutamiini lisamine täiustab selle koostist veelgi.
DMEM, mis on tuletatud Eagle'i minimaalse essentsiaalse keskkonna (EMEM) baasil, pakub võrreldes oma eelkäijaga suuremat aminohapete ja vitamiinide kontsentratsiooni. Seevastu Ham's F-12 põhineb Ham's F-10 keskkonnal, pakkudes täiendavat komplekti olulisi komponente.
Optimaalse rakkude kasvu toetamiseks on tavaline lisada DMEM:Ham's F12-le FBS-i tüüpilises kontsentratsioonis 5–10%. See lisand on vajalik, kuna keskkonnas puuduvad rakkude arenguks olulised kasvuhormoonid, lipiidid ja valgud.
DMEM:Ham's F12 sisaldab pH-puhversüsteemi ja seda täiendatakse sageli fenoolpunasega, mis on pH-indikaator. DMEM:Ham's F12-s või mis tahes bikarbonaatpuhversüsteemi kasutavas keskkonnas kasvatatavad rakud vajavad sobiva pH-taseme säilitamiseks kontrollitud CO2-keskkonda (5–10%).
Kvaliteedikontroll
Steriilselt filtreeritud
Ladustamine ja kõlblikkusaeg
Hoida +2 °C kuni +8 °C juures, valguse eest kaitstult.
Pärast avamist säilitada temperatuuril 4 °C ja kasutada 6–8 nädala jooksul.
Transporditingimused
Ümbritseva õhu temperatuur
Hooldus
Hoida pimedas kohas temperatuuril +2 °C kuni +8 °C. Vältida külmutamist ja sagedast soojendamist temperatuurini +37 °C, kuna see vähendab toote kvaliteeti.
Ärge kuumutage keskkonda üle 37 °C ega kasutage kontrollimatuid soojusallikaid, nagu mikrolaineahjud.
Kui kasutatakse ainult osa keskkonnast, võtke vajalik kogus välja ja soojendage see enne kasutamist toatemperatuurini.
Koostis
Kategooria
Komponendid
Kontsentratsioon (mg/l)
Aminohapped
Glütsiin
18,75
L-alaniin
4,45
L-arginiin HCl
147,50
L-asparagiin H₂O
7,50
L-asparagiinhape
6,65
L-tsüsteiin HCl H₂O
17,56
L-tsüstiin 2 HCl
31,29
L-glutamiinhape
7,35
L-glutamiin
365,00
L-histidiin HCl H₂O
31,48
L-isoleutsiin
54,47
L-leutsiin
59,05
L-lüsiin HCl
91,25
L-metioniin
17,24
L-fenüülalaniin
35,48
L-proliin
17,25
L-seriin
26,25
L-treoniin
53,45
L-trüptofaan
9,02
L-türosiin 2 Na 2 H2O
55,79
L-valiin
52,85
Vitamiinid
D-biotiin
0,0035
Koliinkloriid
8,98
D-kaltsiumpantotenaat
2,24
Foolhape
2,66
Müo-inositool
12,60
Nikotiinamiid
2,02
Püridoksiin-HCl
0,031
Püridoksaal-HCl
2,00
Riboflaviin
0,219
Tiamiin HCl
2,17
B12-vitamiin
0,68
Anorgaanilised soolad
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO3)3 9 H2O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Muud koostisosad
D-glükoos
3151,00
Hüpoksantiin
2,40
HEPES
3574,50
Linoolhape
0,042
Lipoiinhape
0,105
Fenoolpunase naatriumsool
8,63
Putreskiin-2-HCl
0,081
Na-püruvaat
55,00
Tümidiin
0,365
, w: 3,0 g/L glükoos, w: stabiilne glutamiin, w: 2,0 mM naatriumpüruvaat, w: 2,2 g/L NaHCO3")
- 0,02 temperatuuril 20–25 °C. Iga partii on testitud steriilsuse ning mükoplasma ja bakterite puudumise suhtes. Hooldus Hoida külmikus +2 °C kuni +8 °C pimedas kohas. Külmutamine ja soojendamine kuni +37 °C vähendab toote kvaliteeti. Ärge kuumutage keskkonda üle 37 °C ega kasutage kontrollimatuid soojusallikaid (nt mikrolaineahjusid). Kui kasutatakse ainult osa keskkonnast, võtke see kogus pudelist välja ja soojendage toatemperatuuril. Kõigi keskkonna, välja arvatud baaskeskkonna, kõlblikkusaeg on 6–8 nädalat alates avamiskuupäevast. Koostis Komponendid mg/L Anorgaanilised sooladKaltsiumkloriid x 2H2O132,0 Magneesiumsulfaat97,67 Kaaliumkloriid400 Naatriumkloriid6460 Dinatriumvesinikfosfaat (veevaba)504,00 Muud komponendidD(+)-glükoos (veevaba)3000 Glutatioon (redutseeritud)0 Lihapepton600 Fenoolpunane naatriumsool11 AminohappedL-alaniin13,36 L-arginiin x HCl42,14 L-asparagiin x H2O45,03 L-asparagiinhape19,97 L-tsüsteiin x HCl x H2O31,75 L-glutamiin (stabiilne)219,15 L-glutamiinhape22,07 Glütsiin7,51 L-histidiin x HCl x H2O20,96 L-hüdroksüproliin19,67 L-isoleutsiin39,36 L-leutsiin39,36 L-lüsiin x HCl36,54 L-metioniin14,92 L-fenüülalaniin16,52 L-proliin17,27 L-seriin26,28 L-treoniin17,87 L-trüptofaan3,06 L-türosiini dinaatriumsool26,10 L-valiin17,57 Vitamiinidp-aminobensoehape1,0 Askorbiinhape0,56 D(+)-biotiin0,20 D-kaltsiumpantotenaat0,2 Koliinkloriid5 Foolhape10 Müo-inositool36 Nikotinaamid0 Nikotiinhape0,5 Püridoksaal HCl0,5 Püridoksiin HCl0,5 Riboflaviin0,2 Tiamiin HCl0,2 Vitamiin B122,0

Medium 199 pakub mitmesuguseid rakendusi valdkonnas. See võib tõhusalt säilitada kumulus
- ja ootsüütide kompleksi (COC) ja toetada ootsüütide in vitro küpsemist. Lisaks kasutatakse seda aspiratsiooniliinide loputamisel Saksa holsteini lehmadelt munarakkude kogumisel. Lisaks sellele on Medium 199 suurepärane keskkond rottidest saadud südame endoteelirakkude kasvatamiseks. Need rakendused näitavad Medium 199 mitmekülgsust ja kohandatavust erinevate katsete jaoks.
Ajalugu
Medium 199 väljatöötamine 1950. aastatel tähistas olulist edasiminekut koekultuurikeskkondades. Enne selle kasutuselevõttu tuginesid paljud kasvukeskkonnad loomsetele toodetele ja koeekstraktidele. Morgan ja kolleegid tegid aga selles valdkonnas revolutsiooni, sõnastades rakukultuuride jaoks täielikult määratletud toiteallika. Oma katsete kaudu, mis hõlmasid erinevaid vitamiinide, aminohapete ja muude tegurite kombinatsioone, avastasid nad Medium 199 erakordsed kasvu soodustavad omadused.
Kvaliteedikontroll
pH = 7,2 +/
- 0,02 temperatuuril 20-25 °C.
Iga partii on testitud steriilsuse ning mükoplasma ja bakterite puudumise suhtes.
Hooldus
Hoida külmikus +2°C kuni +8°C juures pimedas. Külmutamine ja soojendamine kuni +37° C vähendab toote kvaliteeti.
Ärge kuumutage keskkonda üle 37° C ega kasutage kontrollimatuid soojusallikaid (nt mikrolaineahjusid).
Kui on vaja kasutada ainult osa söötmest, võtke see kogus pudelist välja ja soojendage seda toatemperatuuril.
Kõikide söötmete, välja arvatud põhisöötme säilivusaeg on 8 nädalat alates tootmiskuupäevast.
Koostis
Koostisosad
mg/l
Anorgaanilised soolad
Kaltsiumkloriid x 2H2O
264,92
Raud(III)nitraat x 9H2O
0,72
Magneesiumsulfaat
97,67
Kaaliumkloriid
400,00
Naatriumatsetaat x 3H2O
82,95
Naatriumkloriid
6,800.00
Naatriumdivesinikfosfaat x H2O
140,00
Muud komponendid
Adeniinsulfaat
10,00
AMP
0,20
ATP
1,00
Kolesterool
0,20
2'-Deoksüriboos
0,50
D(+)-glükoosi veevaba
1,000.00
Glutatioon (punane)
0,05
Guaniin x HCl
0,30
Hüpoksantiin
0,30
Fenool punane
10,00
D-Riboos
0,50
Tümiin
0,30
Tween 80
4,90
Uracil
0,30
Ksantiin
0,30
NaHCO3
2,200.00
Aminohapped
L-alaniin
25,00
L-arginiin x HCl
70,00
L-Asparagiinhape
30,00
L-tsüsteiin x HCl x H2O
0,10
L-tsüsteiin
20,00
L-glutamiin stabiilne
149,00
L-glutamiinhape
67,00
Glütsiin
50,00
L-Histidiin x HCl x H2O
21,88
L-hüdroksüproliin
10,00
L-isoleutsiin
20,00
L-leutsiin
60,00
L-Lüsiin x HCl
70,00
L-metioniin
15,00
L-Fenüülalaniin
25,00
L-proliin
40,00
L-Seriin
25,00
L-treoniin
30,00
L-trüptofaan
10,00
L-Türosiin
40,00
L-valiin
25,00
Vitamiinid
4-Aminobensoehape
0,05
Askorbiinhape
0,05
D(+)-Biotiin
0,01
Kaltsiferool
0,10
D-kaltsium-pantotenaat
0,01
Koliinkloriid
0,50
Foolhape
0,01
müo-Inositool
0,05
Menadioon
0,01
Nikotiinhape
0.025
Nikotiinamiid
0.025
Püridoksaal x HCl
0.025
Püridoksool x HCl
0.025
Riboflaviin
0,01
DL-α-tokoferoolfosfaat-dinaatriumsool
0,01
Tiamiin x HCl
0,01
Vitamiin A atsetaat
0,14

IMDM sobib hästi kiiresti paljunevate, suure tihedusega rakukultuuride, sealhulgas Jurkat-, COS-7
- ja makrofaagirakkude kasvatamiseks. Erinevate rakukultuuride jaoks saadaval olevad IMDMi erinevad modifikatsioonid on leitavad meediumivaliku tööriista abil. Vedelkeskkonnad pakuvad olulisi toitaineid kõigi rakukultuuride jaoks. Iga meie kvaliteetne rakukultuurikeskkond on toodetud vastavalt algselt avaldatud valemile või modifikatsioonidele, mis on vajalikud kultuurikeskkonna järjepideva toimimise ja stabiilsuse tagamiseks.
IMDM vs. DMEM
IMDM sisaldab kaaliumnitraati raudnitraadi asemel ning HEPESi ja naatriumpüruvaati. IMDMi lisakomponendid muudavad selle sobivamaks spetsiaalsete rakutüüpide ja spetsiifiliste rakenduste jaoks kui DMEM.
IMDM vs. RPMI
IMDM ja RPMI on erineva koostisega, mis võib olla oluline PMA/ionomütsiini stimuleerimisel. Üks oluline erinevus on Ca2+ kontsentratsioon. Kui RPMI sisaldab 0,42 mM Ca2+, siis IMDM sisaldab 1,49 mM.
Kvaliteedikontroll
pH = 7,2 +/
- 0,02 temperatuuril 20-25 °C.
Iga partii on testitud steriilsuse ning mükoplasma ja bakterite puudumise suhtes.
Hooldus
Hoida külmikus +2°C kuni +8°C juures pimedas. Külmutamine ja soojendamine kuni +37° C vähendab toote kvaliteeti.
Ärge kuumutage keskkonda üle 37° C ega kasutage kontrollimatuid soojusallikaid (nt mikrolaineahjusid).
Kui on vaja kasutada ainult osa söötmest, võtke see kogus pudelist välja ja soojendage seda toatemperatuuril.
Kõikide söötmete, välja arvatud põhisöötme säilivusaeg on 8 nädalat alates tootmiskuupäevast.
Koostis
Koostisosad
mg/l
Anorgaanilised soolad
Kaltsiumkloriid x 2 H2O
219,00
Kaaliumkloriid
330,00
Kaaliumnitraat
0.076
Magneesiumsulfaat veevaba
97,73
Naatriumkloriid
4,505.00
Naatriumdivesinikfosfaat veevaba
109,00
Naatriumseleniit
0,02
Muud komponendid
D(+)-glükoos veevaba
4,500.00
HEPES
5,958.00
Naatriumpüruvaat
110,00
Punane fenool
15,00
Aminohapped
L-alaniin
25,00
L-arginiin x HCl
84,00
L-asparagiin x H2O
25,00
L-asparagiinhape
30,00
L-tsüstiin x 2HCl
91,24
L-Glutamiin
584,00
L-glutamiinhape
75,00
Glütsiin
30,00
L-Histidiin x HCl x H2O
42,00
L-isoleutsiin
104,80
L-leutsiin
104,80
L-Lüsiin x HCl
146,20
L-metioniin
30,00
L-fenüülalaniin
66,00
L-proliin
40,00
L-Seriin
42,00
L-treoniin
95,20
L-trüptofaan
16,00
L-Türosiin x 2Na
104,20
L-Valiin
93,60
Vitamiinid
D(+)-Biootiin
0.013
D-kaltsium-pantotenaat
4,00
Koliinkloriid
4,00
Foolhape
4,00
müo-Inositool
7,20
Rakukultuurikeskkonnad: ülevaade
Eluteaduste valdkonnas on üks olulisemaid meetodeid rakukultuur. Mõiste „rakukultuur” tähendab rakkude, kudede või organite eemaldamist loomalt või taimelt ning nende rakkude, kudede või organite järgnevat siirdamist kunstlikku keskkonda, mis soodustab nende ellujäämist ja/või kasvu. Rakkude optimaalse arengu põhilised keskkonnatingimused on reguleeritud temperatuur, substraat rakkude kinnitumiseks, sobiv kasvukeskkond ning inkubaator, mis hoiab optimaalset pH-taset ja osmolalisust. Rakud vajavad neid tingimusi, et kasvada oma täieliku potentsiaali ulatuses.
Sobiva kasvukeskkonna valik in vitro kasvatamiseks on rakukultuuri protsessis kõige kriitilisem ja olulisem etapp. Kasvukeskkond, mida nimetatakse ka kultuurikeskkonnaks, on vedelik või geel, mis on koostatud soodustama organismide arengut mikroskoopilisel, rakulisel või taimelaadsel tasandil. Rakukultuuri kasvatamiseks kasutatav keskkond sisaldab sageli piisavat energiavarustust ja aineid, mis reguleerivad rakutsüklit. Kultuurikeskkonna peamised koostisosad on aminohapped, vitamiinid, anorgaanilised soolad, glükoos ja seerum. Seerum lisatakse keskkonnale, kuna see toimib kasvufaktorite, hormoonide ja kinnitumisfaktorite allikana. Lisaks toitainete pakkumisele aitab keskkond kaasa ka pH-taseme ja osmolalisuse säilitamisele.
Rakukultuuris kasutatavad keskkonna tüübid
Nii inim- kui ka loomarakke on võimalik kasvatada kas kunstlikus või sünteetilises keskkonnas või täiesti looduslikus keskkonnas, mida on täiendatud looduslike elementidega. Järgnevalt anname ülevaate erinevatest praegu kättesaadavatest keskkonnatüüpidest.
Looduslikud kasvukeskkonnad
Looduslikes kasvukeskkondades leidub ainult looduslikus olekus esinevaid bioloogilisi vedelikke. Looduslikud kasvukeskkonnad on väga kasulikud ja lihtsad mitmesuguste loomarakutüüpide kasvatamiseks. Looduslikke kasvukeskkondi moodustavate täpsete komponentide puudulik mõistmine on peamine tegur, mis mõjutab looduslike kasvukeskkonna kasutamisel saadud tulemuste madalat korratavust.
Kunstlikud kasvukeskkonnad
Kunstlike või sünteetiliste kasvukeskkonna valmistamisel lisatakse toitaineid (nii orgaanilisi kui ka anorgaanilisi), seerumvalke, süsivesikuid, kofaktoreid, vitamiine ja sooli, samuti O₂ ja CO₂ gaasifaase [1].
On välja töötatud mitmesuguseid kunstlikke kasvukeskkondi, et täita ühte või mitut järgmistest funktsioonidest: 1) Vahetu ellujäämine (tasakaalustatud soolalahus täpse pH-väärtuse ja osmootse rõhuga). 2) Pikendatud ellujäämine (tasakaalustatud soolalahus, millele on lisatud erinevaid orgaaniliste kemikaalide ja/või seerumi koostisi). 3) Piiramatu areng. 4) Spetsiifilised funktsioonid.
Kunstlikud kasvukeskkonnad jagunevad neljaks erinevaks klassiks:
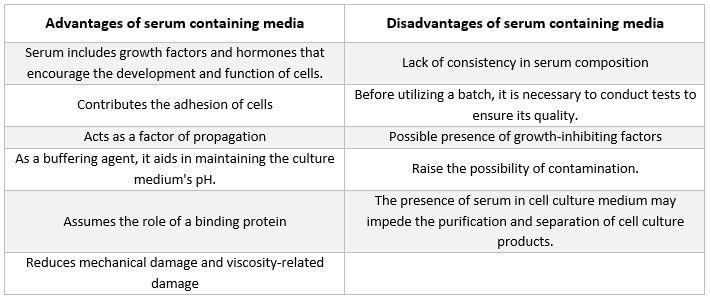
Seerumit sisaldavad keskkonnad
Loomarakkude kasvatamiseks kasutatavates keskkondades leiduvaim lisand on veise loote seerum. Seda lisatakse kultuurikeskkonnale odava lisandina, et saavutada parimad võimalikud kasvutingimused. Lisaks sellele, et seerum toimib ebastabiilsete või vees lahustumatute toitainete, hormoonide ja kasvufaktorite, proteaasi inhibiitorite ning muude ainete transportijana või kelaatorina, seob ja neutraliseerib see ka kahjulikke molekule.
Seerumivaba kasvukeskkond
Seerumi olemasolul keskkonnas on mitmeid puudusi ning see võib põhjustada suuri tõlgendusvigu immunoloogilistes uuringutes [2, 3]. On loodud mitmesuguseid seerumivabasid keskkondi [4, 5]. Need kasvukeskkonnad on üldjuhul spetsiaalselt koostatud ühe rakutüübi kultiveerimiseks, näiteks Thermo Fisher Scientificu tooted „Knockout Serum Replacement“ ja „Knockout DMEM“ ning Stem Cell Technologiesi mTESR-kasvukeskkond [6] tüvirakkude jaoks [7].
Lisaks sisaldavad need keskkonnad kindlaksmääratud koguseid puhastatud kasvufaktoreid, lipoproteiine ja muid valke, mida muidu tavaliselt annab seerum [8]. Neid keskkondi nimetatakse sageli „määratletud kultuurikeskkondadeks”, kuna nende koostisosad on hästi teada.
Keemiliselt määratletud kasvukeskkonnad
Need keskkonnad sisaldavad ülipuhasteid anorgaanilisi ja orgaanilisi komponente, mida ei ole saastanud mingisugune saaste. Need võivad sisaldada ka puhtaid valkude lisandeid, näiteks kasvufaktoreid.
Bakterite või pärmi geneetiline modifitseerimine koos teatud rasvhapete, vitamiinide, kolesterooli ja aminohapete lisamisega viib nende koostisosade tootmiseni [9].
Valguvabad kasvukeskkonnad
Valguvabad kasvukeskkonnad on sellised, mis ei sisalda üldse valku, vaid ainult valguväliseid elemente. Võrreldes seerumilisandiga kasvukeskkondadega soodustab valguvaba keskkonna kasutamine suuremat rakkude paljunemist ja valgu ekspressiooni ning hõlbustab järgnevas protsessis tekkinud toote puhastamist [10–12]. Valgu ei sisaldu sellistes koostistes nagu MEM ja RPMI-1640. Vajaduse korral võib siiski manustada valgu lisandit.
Kasvatuskeskkond ja selle põhikomponendid
Kaubanduslikku kultuurikeskkonna saab osta pulbri või vedeliku kujul ning see sisaldab sageli mitmesuguseid toitaineid, nagu aminohapped, glükoos, soolad, vitamiinid ja muud toidulisandid.
Nende komponentide vajadus on iga rakuliini puhul erinev ning need erinevused on põhjuseks, miks on olemas nii palju erinevaid kasvukeskkonna koostisi. Igal komponendil on oma kindel funktsioon, mida kirjeldatakse järgmistes lõikudes:
Puhversüsteemid
Optimaalsete kasvutingimuste säilitamiseks tuleb reguleerida pH-väärtust, mida tehakse sageli ühe kahest puhverdamissüsteemist:
Looduslik puhverdamissüsteem
CO₂/H₂CO₃ suhe atmosfääris on võrdne sellega, mis on kasvukeskkonnas, luues loodusliku puhvermehhanismi. Selle loodusliku puhvermehhanismi säilitamiseks tuleb kultuure hoida õhukeskkonnas, kus CO₂ sisaldus on 5–10%, mida saavutatakse sageli CO₂-inkubaatori abil. Loodusliku puhvri kasutamise üks suurimaid eeliseid on selle odavus ja ohutus.
HEPES
Zwitterioonil HEPES põhinev keemiline puhverdamine omab suuremat puhverdamisvõimet pH-vahemikus 7,2–7,4 ega vaja reguleeritud gaasikeskkonda. Teatavate rakutüüpide puhul võib suurem HEPES-i annus olla kahjulik. HEPES-i sisaldavad kasvukeskkonnad on samuti palju tundlikumad fluorestsentsvalguse fototoksilise mõju suhtes [13].
Fenoolpunane
pH-indikaator fenoolpunane on sageli lisatud kaubanduslikult kättesaadavatesse kultuurikeskkondadesse, võimaldades pH-väärtuse pidevat jälgimist. Rakude paljunemisel põhjustavad nende poolt toodetud metaboliidid pH-väärtuse muutust ja seega ka keskkonna värvimuutust. Fenoolpunasel on keskkonna värvile kahepoolne mõju: happelise pH juures muutub see kollaseks ja aluselise pH juures lillaks. pH 7,4, mis on rakukultuuri jaoks optimaalne väärtus, muudab keskkonna fluorestsentspunaseks.
Kuid fenoolpunasel on mõned puudused: esiteks suudab fenoolpunane imiteerida mitmete steroidhormoonide, eelkõige östrogeeni toimet [14]. Seetõttu soovitatakse östrogeenitundlike rakkude, nagu rinnakoe, uurimisel kasutada fenoolpunast mittesisaldavat kasvatuskeskkonda. Mitmes seerumivabas koostises häirib fenoolpunase olemasolu naatrium-kaaliumi tasakaalu. Selle mõju võib neutraliseerida, lisades keskkonnale seerumit või veise hüpofüüsi hormooni [15]. Kolmandaks takistab fenoolpunase olemasolu tuvastamist voolutsütomeetrilistes katsetes.
Anorgaanilised soolad
Anorgaanilisi sooli, nagu naatrium-, kaalium- ja kaltsiumioone, sisaldavad kasvukeskkonnad aitavad säilitada osmootilist tasakaalu ja reguleerida membraanipotentsiaali.
Aminohapped
Kuna aminohapped on valkude põhikomponendid, on need oluline osa igast kunagi välja töötatud rakukasvatuskeskkonnast. Kuna rakud ei suuda teatud aminohappeid ise toota, on oluline, et kultuurikeskkonnas oleksid olemas asendamatud aminohapped. Need on vajalikud rakkude paljunemiseks ning nende kontsentratsioon määrab ära maksimaalse saavutatava rakutiheduse. Eriti oluline on asendamatu aminohape L-glutamiin.
L-glutamiin toimib ainevahetuse teisese energiaallikana ja annab lämmastikku NAD-i, NADPH-i ja nukleotiidide tootmiseks. Kuna L-glutamiin on ebastabiilne aminohape, mis aja jooksul muutub vormiks, mida rakud ei suuda ära kasutada, tuleb seda kasvukeskkonnale lisada.
Lisaks võib kasvukeskkonnale lisada mitteolulisi aminohappeid, et asendada kasvu käigus ära kasutatud aminohappeid. Rakke kasvu kiirendatakse ja nende elujõulisust suurendatakse, kui kasvukeskkonnale lisatakse mitteolulisi aminohappeid.
Süsivesikud
Suhkrute kujul esinevad süsivesikud on peamine energiaallikas. Paljudes kasvukeskkondades sisaldub lisaks levinumatele suhkrutele – glükoosile ja galaktoosile – ka maltoosi ja fruktoosi.
Valgud ja peptiidid
Albumiin, transferriin ja fibronektiin on kõige sagedamini kasutatavad valgud ja peptiidid. Need on eriti olulised keskkondades, mis ei sisalda seerumit. Albumiin, transferriin, aprotiniin, fetuiin ja fibronektiin on mõned valgud, mida võib leida seerumist, mis on rikkalik valguallikas.
Albumiin on veres leiduva peamine valk, mille ülesanne on siduda ja transportida erinevaid aineid, sealhulgas vett, sooli, vabu rasvhappeid, hormoone ja vitamiine, erinevate organite ja rakkude vahel. Albumiini võime seonduda kemikaalidega teeb sellest tõhusa vahendi kahjulike ühendite eemaldamiseks rakukultuuri kasvukeskkonnast.
Aprotiniin on rakukultuuri süsteemides kasutatav kaitseaine, kuna see on stabiilne nii neutraalses kui ka happelises pH-keskkonnas ning vastupidav kõrgetele temperatuuridele ja proteolüütiliste ensüümide poolt põhjustatavale lagunemisele. See suudab pärssida mitmeid seriinproteaase, sealhulgas muu hulgas trüpsiini.
Fetuin on glükoproteiin, mida võib loomade loote ja vastsündinute seerumis leida suuremas koguses võrreldes täiskasvanute seerumiga. Lisaks sellele toimib see seriinproteaasi inhibiitorina. Valgu fibronektiin on rakkude adhesiiooni protsessis oluline komponent. Transferriin on valk, mis transportib rauda ja vastutab raua toimetamise eest rakumembraanidesse.
Rasvhapped ja lipiidid
Need mängivad olulist rolli seerumivabas keskkonnas, kui seerum puudub.
Vitamiinid
Rakkude arenguks ja paljunemiseks on vaja arvukaid vitamiine. Rakud ei suuda vitamiine piisavas koguses toota ning seetõttu on need koekultuuris toidulisanditena hädavajalikud.
Rakukultuuris on seerum vitamiinide peamine allikas; samas lisatakse kasvukeskkondadele ka erinevaid vitamiine, et need sobiksid konkreetse rakutüübiga. Kasvu stimuleerimiseks kasutatakse kõige sagedamini B-grupi vitamiine.
Mikroelemendid
Keemilisi elemente, nagu vask, tsink, seleen ja trikarboksüülhappe vaheühendid, nimetatakse mikroelementideks. Mikroelemente lisatakse sageli seerumivabadele kasvukeskkondadele, et asendada neid, mis tavaliselt seerumis esinevad. Need elemendid on olulised keemilised komponendid, mis on vajalikud tervete rakkude arenguks. Paljud biokeemilised reaktsioonid, näiteks ensüümide aktiivsus, sõltuvad teatud mikrotoitainetest.
Kasvukeskkonna lisandid
Teatavatele rakuliinidele soovitatav täielik kasvukeskkond vajab lisakomponente, mida baaskeskkonnas ja seerumis ei ole. Need toidulisandid toetavad rakkude kasvu ja nõuetekohast ainevahetust.
Kuigi hormoonid, kasvufaktorid ja signaalmolekulid on teatud rakuliinide nõuetekohaseks paljunemiseks hädavajalikud, tuleks alati järgida järgmisi ettevaatusabinõusid: kuna lisandite lisamine võib muuta täieliku kasvukeskkonna osmolalisust, mis võib pärssida rakkude arengut, on alati soovitatav kontrollida osmolalisust pärast lisandite lisamist. Enamiku rakuliinide puhul on optimaalne osmolalisus vahemikus 260–320 mOSM/kg.
Antibiootikumid
Antibiootikume kasutatakse sageli bakteriaalsete ja seenhaiguste tekitajate arengu pärssimiseks [16], kuigi need ei ole rakkude kasvu jaoks hädavajalikud. Kuna antibiootikumid võivad varjata mükoplasma ja resistentsete bakterite põhjustatud saastumist, ei soovitata nende rutiinset kasutamist rakukultuurides [17, 18].
Lisaks võivad antibiootikumid häirida ülitundlike rakkude ainevahetust. Sageli kasutatakse MilliporeSigma ja Life Technologiesi toodetud penitsilliini-streptomütsiini kombinatsioone. Plasmocini on kasutatud glioomi rakuliinide TS603, TS516 ja BT260 kultiveerimisel [19] ning see on osutunud efektiivseks mükoplasma saastumise kõrvaldamisel (20).
Seerum
Seerumis leidub albumiine, kasvufaktoreid ja kasvuinhibiitoreid. Seerum on üks rakukultuurikeskkonna olulisemaid komponente, kuna see varustab rakke aminohapete, valkude, vitamiinide (eriti rasvlahustuvate vitamiinidega nagu A, D, E ja K), süsivesikute, lipiidide, hormoonide, kasvufaktorite, mineraalide ja mikroelementidega.
Kultiveeritud rakkude arengu soodustamiseks kasutatakse sageli veiste loote- ja vasikaserumit. Loote seerum on rikkalik kasvufaktorite allikas ning sobib rakkude kloonimiseks ja tundlike rakkude arendamiseks. Vasikaserumit kasutatakse kontaktinhibitsiooni katsetes, kuna selle kasvut soodustav toime on vähenenud. Tavalised kasvukeskkonnad sisaldavad sageli 2–10% seerumit. Seerumi lisamine kultuurikeskkonnale teenib järgmisi eesmärke [21]:
-
Seerum varustab rakke oluliste toitainetega (nii lahuses kui ka valkudega seotuna).
-
Seerumis sisalduvad mitmed kasvufaktorid ja hormoonid, mis osalevad kasvu soodustamises ja spetsiifilises rakutegevuses.
-
See pakub palju seonduvaid valke, nagu albumiin ja transferriin, mis transportivad teisi kemikaale rakku. Näiteks toimetab albumiin rakkudesse rasvu, vitamiine, hormoone jne.
-
Samuti pakub see valke, nagu fibronektiin, mis suurendavad rakkude adhesiooni substraadile. Lisaks toodab see levimiselemente, mis aitavad kaasa rakkude laienemisele enne jagunemist.
-
See toodab proteaasi inhibiitoreid, mis takistavad proteolüüsi rakkudes.
-
See sisaldab ka mineraalaineid, nagu Na+, K+, Zn2+ ja Fe2+.
-
See suurendab kasvatuskeskkonda viskoossust, kaitstes seega rakke mehaaniliste vigastuste eest suspensioonikultuuri segamise ajal.
-
See toimib ka puhvrina.
Viited
[1] Morgan J, Morton H, Parker R. Loomarakkude toitumine koekultuuris; esialgsed uuringud sünteetilise keskkonna kohta. Proc Soc Exp Biol Med. 1950;73:1–8
[2] Kerbel R, Blakeslee D. Loote vasikaserumi komponendi kiire adsorptsioon kultuuris olevate imetajarakkude poolt. Potentsiaalne artefaktide allikas rakuspetsiifiliste antigeenide vastaseerumite uuringutes. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. Seerumi lisamine rakususpensioonide valmistamiseks kasutatavasse keskkonnasse kui võimalik artefaktide allikas rakkude vahendatud reaktsioonide uurimisel põlvelümfisõlme testi abil. J Immunogenet. 1980;7:483–9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Kaubanduslikud seerumivabad kasvukeskkonnad: hübriidoma kasvatamine ja monoklonaalsete antikehade tootmine. J Immunol Methods. 1991;145:175–83
[5] Barnes D, Sato G. Meetodid kultiveeritud rakkude kasvatamiseks seerumivabas keskkonnas. Anal Biochem. 1980;102:255–70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O jt. HSP70 juhib RNA-vaba TDP-43-d anisotroopsetesse tuuma-sisestesse vedelatesse sfäärilistesse kestadesse. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J jt. Downi sündroomist tingitud vananemine häirib närvirakkude eellaste tuuma arhitektuuri. Cell Stem Cell. 2022;29:116–130.e7
[8] Iscove N, Melchers F. Seerumi täielik asendamine albumiini, transferriini ja soja lipiididega lipopolüsahhariidile reageerivate B-lümfotsüütide kultuurides. J Exp Med. 1978;147:923–33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Hübriidoma kasvu ja monoklonaalsete antikehade tootmise jaoks mõeldud keemiliselt määratletud valguvaba keskkonna süstemaatiline täiustamine. J Biotechnol. 1996;45:111–23
[10] Darfler F. Valguvaba kasvukeskkond hübriidomide ja muude immuunsüsteemi rakkude kasvatamiseks. In Vitro Cell Dev Biol. 1990;26:769–78
[11] Barnes D, Sato G. Seerumivaba rakukultuur: ühtne lähenemisviis. Cell. 1980;22:649–55
[12] Hamilton W, Ham R. Hiina hamstri rakuliinide klooniline kasvatamine valguvabas keskkonnas. In Vitro. 1977;13:537–47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Valgusele eksponeeritud HEPES-i sisaldava kultuurikeskkonna tsütotoksiliste mõjude analüüs. In Vitro Cell Dev Biol. 1985;21:282–7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. Fenoolpunane kudekultuurikeskkonnas on nõrk östrogeen: mõju östrogeenile reageerivate rakkude uurimisele kultuuris. Proc Natl Acad Sci U S A. 1986;83:2496–500
[15] Karmiol S. Seerumivabade keskkondade arendamine. In: Master JRW, toim. Animal Cell culture, 3. trükk. Oxford: Oxford University Press; 2000.
[16] Perlman D. Antibiootikumide kasutamine rakukultuurikeskkondades. Methods Enzymol. 1979;58:110–6
[17] McGarrity G. Rakukultuuride mükoplasmainfektsiooni levik ja kontroll. In Vitro. 1976;12:643–8
[18] Masters J, Stacey G. Kasvukeskkonna vahetamine ja rakuliinide passaažimine. „Nat Protoc”. 2007;2:2276–84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M jt. Histooni demetülaas KDM6A tajub otseselt hapnikku, et reguleerida kromatiini ja raku saatust. Science. 2019;363:1217–1222
[20] Molla Kazemiha V, Azari S, Amanzadeh A, Bonakdar S, Shojaei Moghadam M, Habibi Anbouhi M jt. Plasmocin™i tõhusus mitmesugustel mollicutega nakatunud imetajate rakuliinidel võrreldes rakukultuuris tavaliselt kasutatavate antibiootikumidega: kohalik kogemus. Cytotechnology. 2011;63:609–20
[21] Kragh Hansen U. Ligandi seondumise molekulaarsed aspektid seerumalbumiiniga. Pharmacol Rev. 1981;33:17–53