
、L-アラニル-L-グルタミン溶液")
容量:100 ml
保存条件:-15°C以下
無菌性:無菌ろ過済み
安定型グルタミン溶液(L-アラニル-L-グルタミン、200 mM)は、細胞培養培地における従来のL-グルタミンの直接代替として設計された、極めて安定性の高いL-グルタミンのジペプチド形態です。L-グルタミンは必須アミノ酸であり、培養細胞の主要なエネルギー源として、細胞の成長、代謝、タンパク質合成において重要な役割を果たします。
用途と利点
標準的な液体培地では、L-グルタミンは37°Cで比較的速やかに分解され、細胞生存率や実験結果に悪影響を及ぼす可能性のあるアンモニウムイオンなどの有害な副生成物を形成します。安定型グルタミンは、培養条件下で分解されないジペプチド形態を提供することでこの制限を克服します。
細胞は必要な時に酵素的にジペプチド結合を切断しL-グルタミンを放出するため、有害な廃棄物の蓄積を防ぎつつ、継続的な新鮮な供給が保証されます。この特性により、本溶液は長期培養や高密度増殖システムにおいて特に有利です。
製剤と使用方法
L-アラニル-L-グルタミン溶液は、細胞培養グレードの水で200 mMの濃度で調製され、汚染に敏感な用途向けに無菌ろ過されています。実験要件に応じて完全培地に直接希釈できます。製品の安定性を維持するため、-15°C以下で保存し、反復的な凍結融解サイクルを避けてください。
研究用のみ。診断または治療手順での使用は不可。ヒトまたは動物への使用は不可。
")
容量:100 ml
保存条件:+2°C~+8°C
無菌性:無菌ろ過済み
HEPES緩衝液(1 M)、別名N-2-ヒドロキシエチルピペラジン-N-2-エタンスルホン酸は、細胞培養培地で広く使用される両性イオン性有機緩衝剤です。6.7~8.6の生理的範囲で安定したpH条件を維持するように設計されており、in vitro応用時の最適な細胞機能をサポートします。
用途と利点
HEPESは細胞培養システムにおいて、特にCO₂インキュベーター外で細胞を扱う場合に信頼性の高い緩衝能を提供します。培養培地に10~25 mMのHEPESを添加することで、長時間の操作期間中もpH安定性が向上し、一貫した実験条件の維持に貢献します。
この緩衝剤は膜透過性がなく、生化学反応への干渉が最小限で、化学的・酵素的安定性に優れています。これらの特性により、幅広い細胞培養および生化学的用途に適しています。
製剤と使用方法
本品は細胞培養用グレードの水で調製した1 M濃縮液として供給され、汚染に敏感な環境での使用に備え無菌ろ過されています。用途に応じて希望の使用濃度まで希釈可能です。製品の完全性を維持するため、+2°C~+8°Cで保管し、無菌条件下で取り扱ってください。
研究用のみ。診断または治療手順での使用は不可。ヒトまたは動物への使用は不可。
、250 g/L")
容量:50 ml
保存条件:+2°C~+8°C
無菌性:無菌ろ過済み
D-(+)-グルコース溶液(デキストロース溶液)は、細胞代謝の中核成分である天然由来糖D-(+)-グルコースを含む、滅菌済みで即使用可能なサプリメントです。グルコースは、エネルギー産生、糖鎖形成、細胞構造・機能に寄与する糖鎖の形成など、必須の生物学的プロセスに関与しています。
用途と利点
本グルコース溶液は、細胞培養培地における添加剤として、また数多くの細胞生物学・分子生物学応用において広く使用されています。主要な炭素源およびエネルギー源として、グルコースは細胞の成長、増殖、代謝活動をサポートします。生合成経路への関与により、正常な細胞生理機能の維持と実験の一貫性確保にも不可欠です。
製剤と使用方法
本溶液はグルコース250 g/Lの高濃度で供給され、実験要件に応じて培地へ柔軟に希釈可能です。無菌ろ過処理により、汚染に敏感な用途への適性を確保しています。製品品質と安定性を維持するため、+2°C~+8°Cで保管し、無菌的に取り扱ってください。
研究用のみ。診断または治療手順での使用は不可。ヒトまたは動物への使用は不可。
溶液 100倍希釈")
容量:10 ml
保存条件:+2°C ~ +8°C
無菌性:無菌ろ過済み
インスリン・トランスフェリン・セレン(ITS)溶液(100倍希釈)は、幅広い細胞培養用途向けに設計された化学的に定義された添加剤です。主に、低血清または無血清条件下での細胞増殖をサポートするため、基礎細胞培養培地の添加剤として使用されます。
用途と利点
当社のITS補足液は、無血清培地の最適な性能発揮に必要な必須成分を提供します。従来の栄養培地にITSを添加することで、多くの細胞株の日常的な維持に必要な胎児牛血清(FBS)の量を大幅に削減できます。これにより、血清使用に伴う変動を最小限に抑えつつ、一貫した細胞増殖と生存率を維持することが可能になります。
インスリンは主要栄養素の細胞内取り込みと代謝を促進し、トランスフェリンは鉄の輸送を助け、セレンは抗酸化防御と酵素活性に寄与します。これらの成分が相まって、定義された培養システムにおいて、バランスの取れた細胞代謝と再現性の向上を促進します。
製剤および使用方法
インスリン・トランスフェリン・セレン(ITS)は、フェノールレッドを含まないアールズ平衡塩溶液(EBSS)中の100倍濃縮液として供給されます。標準的な用途では、推奨される使用濃度を得るために、適切な基礎培地で1:100に希釈してください。製品の安定性と無菌性を維持するため、+2°C~+8°Cで保存し、無菌条件下で取り扱ってください。
研究用のみ。診断または治療目的での使用はできません。ヒトまたは動物への使用はできません。
")
容量:5 ml
保存条件:+2°C ~ +8°C
無菌性:無菌ろ過済み
ヒト組換えインスリン溶液は、チャイニーズハムスター卵巣(CHO)細胞を含む哺乳類細胞株の培養に一般的に使用される、化学的に定義された培地添加剤です。この細胞培養用グレードの溶液には、Saccharomyces cerevisiaeで発現させた組換えヒトインスリンが含まれており、研究用途において高い純度と安定した性能を保証します。
用途と利点
インスリンは、細胞の増殖と生産性を促進するために、無血清培地や化学的に定義された培地に日常的に添加されます。主要な調節ホルモンとして、インスリンはグルコース、アミノ酸、脂肪酸の細胞内取り込み、利用、および貯蔵をサポートします。また、グリコーゲン、タンパク質、脂質の分解を抑制し、それによって培養システムにおける細胞の生存率と代謝の安定性の向上に寄与します。化学的に定義されたこの製剤は、再現性を高め、繊細な細胞培養ワークフローにおける変動を最小限に抑えます。
生物学的特性と用途
インスリンは、膵島β細胞によって天然に産生される2本鎖のポリペプチドホルモンです。分子量は約5,800 Daです。α鎖とβ鎖は2つの鎖間ジスルフィド結合で連結されており、α鎖には1つの鎖内ジスルフィド結合が含まれています。 細胞培養用途においては、安定性と性能を維持するため、本溶液は無菌条件下で取り扱い、+2°C~+8°Cで保存してください。
研究用のみ。診断または治療目的での使用はできません。ヒトまたは動物への使用はできません。
")
容量:100 ml
保存条件:+2°C~+8°C
無菌性:無菌ろ過済み
ピルビン酸ナトリウム溶液(100 mM)は、細胞培養培地に対して追加の容易に利用可能なエネルギー源を提供する、無菌で即使用可能な添加剤です。ピルビン酸ナトリウムは細胞のエネルギー代謝において重要な役割を果たし、腫瘍細胞などの代謝が活発で急速に増殖する細胞の成長をサポートします。本添加剤は細胞の生存率を高め、培養システムにおける代謝の安定性を維持するのに役立ちます。
用途と利点
本溶液は、ピルビン酸で培地を強化し最適な成長条件を促進するため、日常的な細胞培養で広く使用されています。ATP産生をサポートし、酸化ストレスの軽減に寄与し、培養細胞の代謝パフォーマンス向上に貢献します。細胞培養グレードの水で製造され無菌ろ過されているため、研究ワークフローにおいて一貫した品質と再現性を保証します。
使用方法と互換性
ほとんどの細胞培養用途における推奨最終濃度は1 mMピルビン酸ナトリウムです。100 mMストック溶液を完全培養培地で1:100に希釈することで達成されます。本品は幅広い基礎培地および哺乳類細胞株と互換性があります。製品安定性を維持するため、+2°C~+8°Cで保存し、汚染から保護してください。
研究用のみ。診断または治療手順での使用は不可。ヒトまたは動物での使用は不可。
")
容量:100ml 保存:15℃以下 無菌:無菌ろ過
抗生物質/抗真菌剤溶液(100x)は、細胞培養および関連するラボアプリケーションにおける微生物汚染リスクを低減するために設計された、無菌ですぐに使用できる濃縮液です。この100倍希釈液は、ペニシリン、ストレプトマイシン、アムホテリシンBの定評ある配合で、グラム陽性菌、グラム陰性菌、酵母、糸状菌に対して幅広い抗菌活性を示します。この製剤は、真核細胞培養、細菌培地、およびその他の汚染に敏感なシステムでの使用に適しており、クリーンで一貫性のあるラボ業務をサポートします。
用途と利点 日常的な研究プロトコール用に最適化されたこのソリューションは、細胞培養ワークフローにおける無菌状態の維持に広く使用されています。汚染に敏感な環境で信頼性の高い性能を発揮し、細胞の健康状態や実験の再現性を損なうことなく、微生物の過剰増殖のリスクを低減します。無菌ろ過製剤は、追加の可溶化工程を必要とせず、合理的な培地調製をサポートし、日常的なラボ手順のばらつきを低減します。
使用法と適合性 標準作業濃度を達成するには、溶液を100分の1に希釈し、完全培地に添加します。本製品は、広範な哺乳類細胞株および基礎培地に適合します。安定した在庫により、研究者は信頼できる供給継続性と簡素化されたロジスティクス計画の恩恵を受けることができます。安定性を維持するため、溶液は-15℃以下で保存し、凍結融解の繰り返しから保護してください。 研究用のみ。診断や治療には使用できません。ヒトおよび動物には使用できません。
")
容量:100ml 保存方法+2°C~+8°C 滅菌:無菌ろ過
MEM Non-Essential Amino Acids (100x)は、哺乳動物細胞培養系における細胞の増殖と生存率を高めるために設計された無菌サプリメントです。この製剤は、標準最小必須培地(MEM)に含まれる非必 須アミノ酸を100倍に濃縮したもので、最小限の調製で基礎培地に直接補 充することができます。
応用と利点 このサプリメントは、急速に増殖する細胞や、非必須アミノ酸をデノボで合成する能力を失った細胞株に対して、追加のアミノ酸プールを提供する。生合成の代謝負担を軽減することで、特に栄養に敏感な培養や高密度培養において、成長動態の改善、生存率の延長、実験の一貫性の向上をサポートします。
組成と使用 この溶液は、グリシン、L-アラニン、L-アスパラギン、L-アスパラギン酸、L-グルタミン酸、L-プロリン、L-セリンを含む。MEMおよび他のほとんどの標準培地と互換性がある。使用するには、最終培地に1:100で希釈する。本製品は無菌濾過されており、追加の操作ステップなしですぐに使用できます。 研究用のみ。診断や治療には使用できません。ヒトおよび動物には使用できません。

Accutaseは、標準的な組織培養用プラスチック器具や接着コーティングされた表面から接着性哺乳類細胞を分離するために、トリプシン/EDTAに代わる穏やかな代替品として設計された、即用型の無菌ろ過済み細胞剥離液です。バランスのとれた塩溶液中にタンパク質分解酵素およびコラーゲン分解酵素の活性を組み合わせることで、効果的かつ制御された細胞分離を実現し、細胞表面タンパク質を保持するとともに、継代後の高い生存率と迅速な再付着をサポートします。
Accutaseの配合は、EDTAおよびpHの可視指標としてフェノールレッドを加えたダルベッコリン酸緩衝生理食塩水(DPBS)を基にしています。 酵素は非哺乳類および非細菌由来であるため、Accutaseは幹細胞研究、ワクチン開発ワークフロー、および動物由来または微生物由来の汚染物質を最小限に抑える必要があるあらゆる用途に特に適しています。本溶液は37°Cで自己阻害するため、剥離後に中和試薬や血清含有培地を必要とせず、細胞を新鮮な培地に直接移すことができます。
主な特徴
希釈や再構成が不要な、すぐに使用可能な1倍希釈の滅菌ろ過済み液
穏やかな解離を実現する、タンパク質分解酵素とコラーゲン分解酵素の複合活性
ロット間の均一性を確保するため、各バッチの解離活性は規定値に標準化されています
非哺乳類および非細菌由来の酵素
37 °Cで自己阻害するため、中和溶液は不要
EDTAを含むダルベッコPBSで調製
視覚的pH指示薬としてフェノールレッドを含有
pH 6.8~7.8
代表的な用途
Accutase は、ヒト胚性幹細胞 (hESC)、ヒト誘導多能性幹細胞 (iPSC)、神経幹細胞、初代神経細胞、および HeLa、HEK 293、CHO、MDCK、Vero、NIH/3T3、BHK-21、A549 などの日常的に培養される接着性細胞株など、多種多様な接着性および感受性の高い細胞を穏やかに解離します。 代表的な用途には以下が含まれます:
接着性哺乳類細胞の日常的な継代およびパッシング
hESC、iPS細胞、およびその他の感受性の高い細胞株の穏やかな単一細胞分離
フローサイトメトリーおよびFACS解析のためのサンプル調製
エピトープの完全性が重要な細胞表面マーカーの解析
細胞遊走、増殖、アポトーシスアッセイ
血清飢餓法による静止期アッセイおよび癌遺伝子導入研究
腫瘍細胞および神経堤細胞の遊走アッセイ
バイオリアクターワークフローにおける生産スケールアップ
日常的な作業では、培養表面75 cm²あたり約10 mlのAccutaseを添加し、室温で5~10分間インキュベートします。最適なインキュベーション時間は各細胞株ごとに決定する必要があり、1時間を超えてはいけません。添加前に、残留血清および二価陽イオンを除去するため、カルシウムおよびマグネシウムを含まないDPBSなどのCa²⁺/Mg²⁺フリー塩溶液で細胞層を洗浄してください。
取り扱いおよび保存
未開封のボトルは、-15 °C以下で凍結保存してください。解凍は室温、または+2 °C~+8 °Cで一晩かけて行ってください。高温は酵素活性を低下させるため、37 °Cの水浴でAccutaseを解凍しないでください。解凍後、溶液は+2 °C~+8 °Cで最長2ヶ月間保存可能です。室温での保存は避けてください。 使用前に試薬を37 °Cまで予温しないでください。室温で洗浄済みの細胞に直接添加してください。長期保存のためには、繰り返しの解凍サイクルを避けるため、単回使用分への分注が推奨されます。常に無菌条件下で作業してください。
品質
厳格な品質基準に基づいて製造されています。Accutaseの各バッチは、無菌ろ過され、無菌性、pH、外観、および解離活性について試験されており、ロット間の性能の一貫性と再現性が確保されています。
製品仕様
仕様
詳細
製品タイプ細胞剥離/解離試薬
形態滅菌ろ過済み液体、即用型
容量100 ml
使用濃度1x(即用型)
酵素活性タンパク質分解およびコラーゲン分解の複合作用
酵素の由来非哺乳類および非細菌由来
緩衝液系EDTA含有ダルベッコPBS
pH指示薬フェノールレッド
pH範囲6.8 – 7.8
外観透明、淡紅色~橙色の溶液
保存温度-15 °C以下
解凍後の安定性+2 °C~+8 °Cで最大2ヶ月
推奨使用量培養面積75 cm²あたり約10 ml
標準的な培養時間室温で5~10分
輸送条件ドライアイスで凍結
使用目的研究用および製造用のみ
製剤(1リットルあたりの組成)
成分
濃度 (mg/L)
無機塩
塩化ナトリウム(NaCl)8000.00
リン酸水素二ナトリウム (Na2HPO4)1150.00
塩化カリウム (KCl)200.00
リン酸二水素カリウム (KH₂PO₄)200.00
その他の成分
EDTA・4Na(テトラナトリウムEDTA)220.00
フェノールレッド3.00
独自配合酵素ブレンド(タンパク質分解活性およびコラーゲン分解活性)1x
AccutaseはInnovative Cell Technologies, Inc.の登録商標です。
、w:2mM L-グルタミン、w:2.2g/L NaHCO3、w:EBSS")
この即用型の滅菌ろ過済み液体培地には、アール平衡塩溶液(EBSS)、2 mM L-グルタミン、D-グルコース(1.0 g/L)、および2.2 g/Lの炭酸水素ナトリウム(NaHCO3)が添加されており、CO2制御インキュベーター環境(通常5 % CO2)での使用に適しています。配合されているフェノールレッドはpH指示薬として機能し、細胞培養中の培地状態を視覚的に簡単に確認することができます。
主な特徴
Earle’s Balanced Salt Solution(EBSS)を含む、従来のイーグルMEM配合
2 mM L-グルタミン含有 – すぐに使用可能
2.2 g/L の重炭酸ナトリウム – 5 % CO2 インキュベーション用に緩衝調整済み
主炭素源としてD-グルコース(1.0 g/L)を含有
pH指示薬としてフェノールレッドを含有
HEPESおよびピルビン酸ナトリウムは含まれていません
滅菌ろ過済みの液体培地、すぐに使用可能
pH 7.0 – 7.6
代表的な用途
EMEMは、HeLa、HEK 293、Vero、MRC-5、L-929、BHK-21、および多くの初代細胞を含む、多種多様な哺乳類細胞株の培養をサポートします。一般的な用途には以下が含まれます:
付着性細胞株の日常的な維持および増殖
ウイルスの増殖およびワクチン製造ワークフロー
細胞毒性試験およびバイオアッセイ
トランスフェクションおよびタンパク質発現研究
細胞生物学および分子生物学における基礎研究
最適な細胞増殖のため、EMEMには通常、5~10 % の胎児牛血清(FBS)が添加され、細胞株によっては、非必須アミノ酸(NEAA)やペニシリン/ストレプトマイシンなどの抗生物質も添加されます。
取り扱いおよび保存
未開封のボトルは、光を避けて+2°C~+8°Cで保存してください。開封後は無菌条件下で使用してください。溶液中のL-グルタミンは徐々に分解されます。最適な性能を得るためには、開封後4週間以内に培地を使用するか、長期保存する場合は使用前に新鮮なL-グルタミンを補充することを推奨します。細胞に添加する前に、培地を37°Cまで温めてください。
品質
厳格な品質基準に基づいて製造されています。各バッチは、細胞培養用途において一貫した性能を確保するため、無菌性、pH、浸透圧、およびエンドトキシンレベルについて検査されています。
製品仕様
仕様
詳細
製品タイプMEM
製品カテゴリ細胞培養培地
形態液体
滅菌はい
容量500 ml
L-グルタミンL-グルタミン含有 (2 mM)
グルコースグルコース配合 (1.0 g/L)
重炭酸ナトリウムNaHCO3 含有 (2.2 g/L)
HEPESHEPESなし
ピルビン酸ナトリウムピルビン酸ナトリウムなし
フェノールレッドフェノールレッドを含む
塩溶液アール平衡塩溶液(EBSS)
pH7.0 – 7.6
エンドトキシン含有量未指定
保存+2 °C ~ +8 °C
製剤(1リットルあたりの組成)
成分
濃度 (mg/L)
無機塩
塩化カルシウム・2H2O265.00
硫酸マグネシウム97.72
塩化カリウム400.00
塩化ナトリウム6,800.00
無水リン酸二水素ナトリウム122.00
重炭酸ナトリウム (NaHCO3)2,200.00
アミノ酸
L-アルギニン・塩酸塩126.00
L-シスチン・2HCl31.30
L-グルタミン292.00
L-ヒスチジン・塩酸・水42.00
L-イソロイシン52.00
L-ロイシン52.00
L-リジン・塩酸塩72.50
L-メチオニン15.00
L-フェニルアラニン32.00
L-スレオニン48.00
L-トリプトファン10.00
L-チロシン・2Na・2H2O51.90
L-バリン46.00
ビタミン
D-パントテン酸カルシウム1.00
塩化コリン1.00
葉酸1.00
ミオイノシトール2.00
ニコチンアミド1.00
ピリドキサール・塩酸塩1.00
リボフラビン0.10
チアミン・塩酸塩1.00
その他の成分
D(+)-グルコース1,000.00
フェノールレッド10.00

Freeze Medium CM-1の主な特徴は以下の通り:
幅広い適合性:プライマリー細胞、幹細胞、樹立細胞株など、幅広い種類の細胞に有効。
高い生存率融解後の細胞の回復と生存率を最大化するように最適化されており、信頼性の高い実験結果を保証する。
すぐに使えるすぐに使用できるように調製・滅菌されているため、調製時間とコンタミネーションのリスクが軽減される。
安定性の向上:標準的な凍結保存条件下で一貫した性能を維持し、再現性のある結果を保証します。
長い保存期間:CM-1は血清を含む、すぐに使用可能な凍結保存培地で、冷蔵庫で最長1年間保存可能です。
細胞の凍結にCM-1を使用する
接着細胞および浮遊細胞の凍結にCM-1を使用するには、以下の手順に従う:
接着細胞の場合は、洗浄して培養基材から剥離する。浮遊細胞の場合は、そのまま次のステップに進む。
細胞を数え、適切な濃度であることを確認する。
細胞を遠心してペレット化し、CM-1凍結培地に再懸濁する。
再懸濁した細胞をクライオバイアルに移す。
細胞を長期保存に移す前に、緩慢凍結法を用いる。
方法
手順
手順
❄️
マニュアル凍結
段階的に温度を下げ、細胞の生存性を確保する方法。
1️⃣ 細胞を凍結培地中、4℃のフリーザーに40分間置く。
2️ ⃣ -80℃のフリーザーに24時間移す。
3️ ⃣ 長期保存のため、細胞を液体窒素で保存する。
❄️
Mr.Frostyの使用
電力を使わずに凍結速度を制御できる便利な装置。
1️ ⃣ 細胞を凍結培地を入れたクライオバイアルに準備する。
2️ ⃣ Mr.Frosty容器にクライオバイアルを入れる。
3️⃣ 液体窒素に移す前に、-80℃で24時間保存する。
❄️
コントロールレートフリーザー
サーモ・フィッシャー社などの高精度フリーザーで、温度を 制御して下げるように設計されている。
1️ ⃣ 徐々に温度が下がるように装置をプログラムする。
2️ ⃣ 準備した細胞をフリーザーに入れる。
3️ ⃣ 凍結サイクル終了後、細胞を液体窒素に移す。
クライオバイアルは-130℃以下で保存するか、長期保存のために液体窒素中で保存する。
成分
FBS、DMSO、グルコース、塩類を含む。
緩衝能:pH=7.2~7.6
Cytion's Freeze Medium CM-1 は、凍結保存のための信頼性の高いソリューションを提供し、幅広い研究用途において解凍後の高い細胞生存率と機能性を保証します。

Ham's F-12K(Kaighn's)培地は、細胞培養条件を最適化するために注意深く調合されている。アミノ酸やピルビン酸ナトリウムのような必須成分や、プトレシン、チミジン、ヒポキサンチン、亜鉛などの追加元素の濃縮組成が特徴である。このような添加により、研究者は培地に最小限の血清や特定の細胞タイプ用に定義された成分を補充することができ、正確な実験条件を容易にすることができる。
注目すべきは、ハムのF-12K(カイグン)培地にはタンパク質や成長因子が含まれていないことである。その結果、成長因子やウシ胎児血清(FBS)の補充が必要となることが多く、研究者は培地を特定の細胞株の要求に合わせて調整することができる。最適なパフォーマンスを得るためには、FBSの濃度を細胞株ごとに注意深く最適化し、最適な増殖と機能性を確保する必要がある。
生理的pHを維持するため、Ham's F-12K(Kaighn's)培地は炭酸水素ナトリウム緩衝系(2.5g/L)を採用しており、培養中は5-10%のCO2環境にコントロールする必要がある。これにより、培地のpHは細胞の成長と生存に理想的な範囲に保たれます。
品質管理
pH = 7.2 +/
- 0.02 (20-25℃)。
各ロットは無菌検査済みで、マイコプラズマやバクテリアは検出されていない。
メンテナンス
冷蔵保存(+2℃~+8℃、暗所)。凍結および+37℃までの加温は、製品の品質を最小限に抑えます。
培地を37℃以上に加熱したり、制御不能な熱源(電子レンジなど)を使用しないでください。
メデュームの一部だけを使用する場合は、その分をボトルから取り出し、室温で温めてください。
基本培地以外の培地の保存期間は、製造日から8週間です。
組成
成分
mg/L
無機塩類
塩化カルシウム×2H2O
135,24
硫酸銅(II)×5H2O
0,00
硫酸鉄(II)×7H2O
0,83
塩化マグネシウム×6H2O
105,72
硫酸マグネシウム×7H2O
394,49
塩化カリウム
283,29
リン酸二水素カリウム
58,52
塩化ナトリウム
7597,20
リン酸水素二ナトリウム無水
115,02
硫酸亜鉛×7H2O
0,14
その他の成分
無水D(+)-グルコース
1260,00
ヒポキサンチン
4,08
DL-α-リポ酸
0,21
フェノールレッド
3,00
プトレシン×2塩酸
0,32
ピルビン酸ナトリウム
220,00
NaHCO3
2500,00
チミジン
0,73
アミノ酸
L-アラニン
17,82
L-アルギニン×塩酸塩
421,40
L-アスパラギン×H2O
30,02
L-アスパラギン酸
26,62
L-システイン x HCl x H2O
70,24
L-グルタミン
292,20
L-グルタミン酸
29,42
グリシン
15,01
L-ヒスチジン×HCl×H2O
41,92
L-イソロイシン
7,87
L-ロイシン
26,24
L-リジン×HCl
73,04
L-メチオニン
8,95
L-フェニルアラニン
9,91
L-プロリン
69,06
L-セリン
21,02
L-スレオニン
23,82
L-トリプトファン
4,08
L-チロシン
10,87
L-バリン
23,42
ビタミン
D(+)-ビオチン
0,07
D-パントテン酸カルシウム
0,48
塩化コリン
13,96
葉酸
1,32
ミオ-イノシトール
18,02
ニコチンアミド
0,04
ピリドキシンx塩酸塩
0,06
リボフラビン
0,04
チアミンx塩酸塩
0,34
ビタミンB12
1,36

リン酸緩衝生理食塩水(PBS)は、生物学および化学の研究において広く使用されている緩衝液である。組織処理や細胞培養を含む様々な実験手順において、pHバランスと浸透圧の維持に重要な役割を果たしています。当社のPBS溶液は、あらゆる実験において安定性と信頼性を確保するために、高純度成分を用いて細心の注意を払って調合されています。当社のPBSの浸透圧とイオン濃度は、人体の浸透圧に近いため、ほとんどの細胞に対して等張で無毒です。
PBS溶液の組成
当社のPBS溶液は、超高純度グレードのリン酸緩衝液と生理食塩水をpH調整したものです。1倍濃度で、以下を含む:
8000 mg/L 塩化ナトリウム(NaCl)
200 mg/L 塩化カリウム(KCl)
1150 mg/L 二塩基性無水リン酸ナトリウム (Na2HPO4)
200 mg/L 無水リン酸一塩基性カリウム (KH2PO4)
この組成により、最適なpHとイオンバランスが確保され、幅広い生物学的用途に適しています。
PBS溶液の用途
当社のPBS溶液は、生物学研究における様々な用途に最適です。等張で無毒性であるため、物質の希釈や細胞容器の洗浄に適しています。EDTAを含むPBS溶液は、付着・凝集した細胞の剥離に効果的です。しかし、亜鉛のような二価の金属は沈殿の原因となるため、PBSに添加すべきではない。このような場合は、Good's緩衝液を推奨する。さらに、当社のPBS溶液は、SARS-CoV-2を含むRNAウイルスの輸送および保存のためのウイルス輸送培地の代替品として許容される。
品質管理
無菌ろ過
保管および保存期間
遮光し、+2℃~+25℃で保存する。
開封後は2℃~25℃で保存し、24ヶ月以内に使用してください。
出荷条件
周囲温度
メンテナンス
冷蔵で+2℃~+8℃の暗所に保管する。製品の品質を低下させるため、凍結や+37℃への頻繁な加温は避けてください。
培地を37℃以上に加熱したり、電子レンジのような制御されていない熱源を使用しないでください。
培地の一部のみを使用する場合は、必要な量を取り出し、室温まで温めてから使用してください。
組成
カテゴリー
成分
濃度(mg/L)
塩類
塩化カリウム
200
無水リン酸一塩基性カリウム
200
塩化ナトリウム
8000
無水リン酸二塩基性ナトリウム
1150

RPMI1640培地は、当初ヒト白血病細胞の懸濁培養と単層培養の両方をサポートするために設計されたが、研究者や市販の供給業者による改良を経て、多様な哺乳動物細胞に適した培地へと進化した。HeLa、Jurkat、MCF-7、PC12、PBMC、アストロサイト、癌腫などの細胞株との相性は抜群である。
RPMI1640培地は、そのユニークな組成により、他の細胞培養培地とは一線を画している。かなりの量のリン酸、アミノ酸、ビタミンを含んでいる。特に、イーグル最小必須培地やダルベッコ変法イーグル培地には含まれていないビオチン、ビタミンB12、PABAが含まれている。さらに、RPMI1640培地は、ビタミン・イノシトールとコリンの濃度が著しく高い。しかし、タンパク質、脂質、成長因子は含まれていない。従って、細胞増殖に最適な条件を提供するためには、10%ウシ胎児血清(FBS)の補充が一般的に必要となる。
RPMI1640培地の緩衝系は炭酸水素ナトリウムに依存しており、生理学的に適切なpHを維持するためには5-10%のCO2環境が必要である。還元剤であるグルタチオンが含まれていることが、この培地と他の培地との違いをさらに際立たせている。
品質管理
無菌ろ過
保存と賞味期限
遮光し、+2℃~+8℃で保存する。
開封後は4℃で保存し、6~8週間以内にご使用ください。
出荷条件
周囲温度
メンテナンス
冷蔵で+2℃~+8℃の暗所に保管。凍結や+37℃までの頻繁な加温は、製品の品質を低下させるので避けてください。
培地を37℃以上に加熱したり、電子レンジのような制御されていない熱源を使用しないでください。
培地の一部のみを使用する場合は、必要な量を取り出し、室温まで温めてから使用してください。
組成
カテゴリー
成分
濃度 (mg/L)
アミノ酸
グリシン
10.00
L-アラニル-L-グルタミン
434.40
L-アルギニン
200.00
L-アスパラギンH2O
56.82
L-アスパラギン酸
20.00
L-シスチン2塩酸塩
65.20
L-グルタミン酸
20.00
L-ヒスチジンHClH2O
20.27
L-ヒドロキシ-L-プロリン
20.00
L-イソロイシン
50.00
L-ロイシン
50.00
L-リジン塩酸塩
40.00
L-Methionine
15.00
L-Phenylalanine
15.00
L-Proline
20.00
L-Serine
30.00
L-Threonine
20.00
L-Tryptophan
5.00
L-チロシン2Na2H2O
28.83
L-バリン
20.00
ビタミン
p-アミノ安息香酸
1.00
D-ビオチン
0.20
塩化コリン
3.00
パントテン酸カルシウム
0.25
葉酸
1.00
ミオイノシトール
35.00
ニコチンアミド
1.00
600%0000 ピリミホスメチル
1.00
600%0000 ピリミホスメチル
0.20
600%0000 ピリミホスメチル
1.00
ビタミンB12
0.005
無機塩類
Ca(NO3)2 4H2O
100.00
塩化カリウム
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
その他の成分
D-グルコース
2000.00
還元L-グルタチオン
1.00
フェノールレッドナトリウム塩
5.30
、w: 3.1 g/Lグルコース、w: 2.5 mM L-グルタミン、w: 15 mM HEPES、w: 0.5 mMピルビン酸ナトリウム、w: 1.2 g/L NaHCO3")
この独自の配合は、ダルベッコ改良イーグル培地(DMEM)とハムF-12(ハム栄養混合液F-12)を正確な1:1の比率で組み合わせたものです。L-グルタミンの添加により、その組成はさらに強化されています。
イーグル最小必須培地(EMEM)を起源とするDMEMは、その前身と比較してアミノ酸およびビタミンの濃度が高くなっています。一方、ハムF-12はハムF-10培地を基にしており、必須成分の補完的なセットを提供します。
最適な細胞増殖をサポートするため、DMEM:ハムズF-12に通常5~10%濃度のFBSを添加するのが一般的です。この添加は、培地に細胞の発達に不可欠な成長ホルモン、脂質、タンパク質が不足しているため必要です。
DMEM:ハムズF-12にはpH緩衝システムが組み込まれており、pH指示薬であるフェノールレッドが添加されることが多い。DMEM:ハムズF-12、あるいは重炭酸塩緩衝システムを利用するあらゆる培地で培養される細胞は、適切なpHレベルを維持するために、5~10%のCO2濃度に制御された環境を必要とする。
品質管理
滅菌ろ過済み
保存および有効期限
+2°C~+8°Cで、光を避けて保存してください。
開封後は 4°C で保存し、6~8 週間以内に使用してください。
輸送条件
周囲温度
保管
+2°C~+8°Cの暗所で冷蔵保存してください。製品の品質が低下するため、凍結や +37°C への頻繁な加熱は避けてください。
培地を 37°C 以上に加熱したり、電子レンジなどの制御できない熱源を使用したりしないでください。
培地の一部のみを使用する場合は、必要な量を取り出し、使用前に室温まで温めてください。
成分
カテゴリー
成分
濃度 (mg/L)
アミノ酸
グリシン
18.75
L-アラニン
4.45
L-アルギニン塩酸塩
147.50
L-アスパラギン・水和物
7.50
L-アスパラギン酸
6.65
L-システイン塩酸塩・水和物
17.56
L-シスチン・2HCl
31.29
L-グルタミン酸
7.35
L-グルタミン
365.00
L-ヒスチジン塩酸塩・水和物
31.48
L-イソロイシン
54.47
L-ロイシン
59.05
L-リジン塩酸塩
91.25
L-メチオニン
17.24
L-フェニルアラニン
35.48
L-プロリン
17.25
L-セリン
26.25
L-トレオニン
53.45
L-トリプトファン
9.02
L-チロシン 2Na・2H₂O
55.79
L-バリン
52.85
ビタミン
D-ビオチン
0.0035
塩化コリン
8.98
D-パントテン酸カルシウム
2.24
葉酸
2.66
ミオイノシトール
12.60
ニコチンアミド
2.02
ピリドキシン塩酸塩
0.031
ピリドキサール塩酸塩
2.00
リボフラビン
0.219
チアミン塩酸塩
2.17
ビタミンB12
0.68
無機塩
CaCl2・2H2O
154.50
硫酸銅・五水和物
0.0013
Fe(NO₃)₃・9H₂O
0.05
FeSO4・7H2O
0.417
KCl
311.80
MgCl2・6H2O
61.20
MgSO4・7H2O
100.00
NaCl
6996.00
NaHCO3
1200.00
Na₂HPO₄
71.02
NaH₂PO₄・2H₂O
70.87
ZnSO4・7H2O
0.432
その他の成分
D-グルコース
3151.00
ヒポキサンチン
2.40
HEPES
3574.50
リノール酸
0.042
リポ酸
0.105
フェノールレッドナトリウム塩
8.63
プトレシン2塩酸塩
0.081
ピルビン酸ナトリウム
55.00
チミジン
0.365
、w: 3.0 g/L グルコース、w: 安定グルタミン、w: 2.0 mM ピルビン酸ナトリウム、w: 2.2 g/L NaHCO3")

199培地は、様々な分野での応用が可能です。卵丘細胞-卵母細胞複合体(COC)を効果的に維持し、卵子の体外成熟をサポートすることができる。さらに、ドイツのホルスタイン牛から卵子を採取する際の吸引ラインの洗浄にも使用されています。さらに、199培地はラット由来の心臓内皮細胞の培養に最適な培地です。これらの用途は、199培地の多様性と様々な実験ニーズへの適応性を示している。
歴史
1950年代に199培地が開発されたことは、組織培養培地における重要な進歩であった。この培地の登場以前は、多くの培地が動物由来の製品や組織抽出物に依存していた。しかし、モーガンらは細胞培養のために完全に定義された栄養源を処方することにより、この分野に革命を起こした。ビタミン、アミノ酸、その他の因子の様々な組み合わせを含む実験を通して、彼らは199培地の卓越した成長促進特性を発見したのである。
品質管理
pH=7.2+/-0.02、20-25℃。
各ロットは、マイコプラズマとバクテリアの無菌検査済み。
メンテナンス
冷蔵保存(+2℃~+8℃、暗所)。凍結および+37℃までの加温は、製品の品質を最小限に抑える。
培地を37℃以上に加熱したり、制御不能な熱源(電子レンジなど)を使用しないでください。
メデュームの一部のみを使用する場合は、その分をボトルから取り出し、室温で温めてください。
基本培地以外の培地の賞味期限は、製造日から8週間です。
組成
成分
mg/L
無機塩類
塩化カルシウム×2H2O
264,92
硝酸鉄(III)×9H2O
0,72
硫酸マグネシウム
97,67
塩化カリウム
400,00
酢酸ナトリウム×3H2O
82,95
塩化ナトリウム
6,800.00
リン酸二水素ナトリウム×H2O
140,00
その他の成分
アデニン硫酸塩
10,00
AMP
0,20
ATP
1,00
コレステロール
0,20
2'-デオキシリボース
0,50
無水D(+)-グルコース
1,000.00
グルタチオン(赤)
0,05
グアニン×塩酸
0,30
ヒポキサンチン
0,30
フェノールレッド
10,00
D-リボース
0,50
チミン
0,30
ツイーン80
4,90
ウラシル
0,30
キサンチン
0,30
NaHCO3
2,200.00
アミノ酸
L-アラニン
25,00
L-アルギニン x HCl
70,00
L-アスパラギン酸
30,00
L-システイン x HCl x H2O
0,10
L-シスチン
20,00
L-グルタミン安定
149,00
L-グルタミン酸
67,00
グリシン
50,00
L-ヒスチジン×HCl×H2O
21,88
L-ヒドロキシプロリン
10,00
L-イソロイシン
20,00
L-ロイシン
60,00
L-リジン x HCl
70,00
L-メチオニン
15,00
L-フェニルアラニン
25,00
L-プロリン
40,00
L-セリン
25,00
L-スレオニン
30,00
L-トリプトファン
10,00
L-チロシン
40,00
L-バリン
25,00
ビタミン
4-アミノ安息香酸
0,05
アスコルビン酸
0,05
D(+)-ビオチン
0,01
カルシフェロール
0,10
D-パントテン酸カルシウム
0,01
塩化コリン
0,50
葉酸
0,01
ミオイノシトール
0,05
メナジオン
0,01
ニコチン酸
0.025
ニコチンアミド
0.025
ピリドキサール塩酸塩
0.025
ピリドキサール×塩酸塩
0.025
リボフラビン
0,01
DL-α-トコフェロールリン酸二ナトリウム塩
0,01
チアミン×塩酸塩
0,01
酢酸ビタミンA
0,14

IMDMは、Jurkat、COS-7、マクロファージ細胞など、急速に増殖する高密度細胞培養に適しています。様々な細胞培養用途に利用可能なIMDMの様々な変更は、培地選択ツールを使用して見つけることができます。液体培地は、あらゆる細胞培養アプリケーションに不可欠な栄養素を提供します。当社の高品質細胞培養培地は、培地の安定した性能と安定性のために必要な、最初に公表された処方または変更に従って製造されています。
IMDMとDMEMの比較
IMDMは、硝酸第二鉄、HEPES、ピルビン酸ナトリウムの代わりに硝酸カリウムを含んでいます。IMDMの追加成分により、DMEMよりも特殊な細胞種や特定の用途に適しています。
IMDMとRPMIの比較
IMDMとRPMIは、PMA/イオノマイシン刺激に関連すると思われる配合が異なる。重要な違いの一つは、Ca2+の濃度である。RPMIが0.42mMのCa2+を含むのに対し、IMDMは1.49mMを含む。
品質管理
pH = 7.2 +/
- 0.02 at 20-25℃。
各ロットは、無菌性、マイコプラズマおよびバクテリアの非存在について検査済み。
保存方法
冷蔵保存(+2℃~+8℃、暗所)。凍結および+37℃までの加温は、製品の品質を最小限に抑える。
培地を37℃以上に加熱したり、制御不能な熱源(電子レンジなど)を使用しないでください。
メデュームの一部のみを使用する場合は、その分をボトルから取り出し、室温で温めてください。
基本培地以外の培地の賞味期限は、製造日から8週間です。
組成
成分
mg/L
無機塩類
塩化カルシウム×2 H2O
219,00
塩化カリウム
330,00
硝酸カリウム
0.076
無水硫酸マグネシウム
97,73
塩化ナトリウム
4,505.00
無水リン酸二水素ナトリウム
109,00
亜セレン酸ナトリウム
0,02
その他の成分
無水D(+)-グルコース
4,500.00
HEPES
5,958.00
ピルビン酸ナトリウム
110,00
フェノールレッド
15,00
アミノ酸
L-アラニン
25,00
L-アルギニン塩酸塩
84,00
L-アスパラギン×H2O
25,00
L-アスパラギン酸
30,00
L-シスチン×2HCl
91,24
L-グルタミン
584,00
L-グルタミン酸
75,00
グリシン
30,00
L-ヒスチジン×HCl×H2O
42,00
L-イソロイシン
104,80
L-ロイシン
104,80
L-リジン x HCl
146,20
L-メチオニン
30,00
L-フェニルアラニン
66,00
L-プロリン
40,00
L-セリン
42,00
L-スレオニン
95,20
L-トリプトファン
16,00
L-チロシン×2Na
104,20
L-バリン
93,60
ビタミン
D(+)-ビオチン
0.013
D-パントテン酸カルシウム
4,00
塩化コリン
4,00
葉酸
4,00
ミオイノシトール
7,20
細胞培養培地:概要
生命科学の分野において、最も重要な手法の一つが細胞培養である。「細胞培養」とは、動物や植物から細胞、組織、あるいは臓器を取り出し、それらの生存および/または増殖に適した人工環境に移植することを指す。 細胞が最適に発育するために必要な基本的な環境条件としては、温度管理、細胞が付着するための基質、適切な培養液、そして最適なpHおよび浸透圧を維持するインキュベーターが挙げられます。細胞がその潜在能力を最大限に発揮して増殖するためには、これらの条件が整っている必要があります。
in vitro培養に適した増殖培地の選択は、細胞培養において最も重要かつ不可欠な段階である。 培養液(カルチャーメディアとも呼ばれる)とは、微生物、細胞、あるいは植物などのレベルで生物の発育を促進するように調製された液体またはゲル状の物質である。細胞の培養に使用される培養液には、多くの場合、十分なエネルギー源と、細胞周期を制御する物質が含まれている。 培養培地の主な成分には、アミノ酸、ビタミン、無機塩、グルコース、および血清が含まれます。血清は、成長因子、ホルモン、および付着因子の供給源として機能するため、培地に添加されます。培地は栄養素を供給するだけでなく、pHや浸透圧レベルの維持にも寄与します。
細胞培養で使用される培地の種類
ヒト細胞も動物細胞も、人工または合成培地、あるいは天然成分を添加した完全な天然培地のいずれでも培養することが可能です。以下では、現在利用可能なさまざまな培地の種類について概要を説明します。
天然培地
天然培地には、自然の状態にある生体液のみが含まれます。天然培地は、多種多様な動物細胞の培養において非常に有用であり、扱いも容易です。天然培地を構成する正確な成分が十分に解明されていないことが、天然培地を用いた実験結果の再現性が低い主な要因となっています。
人工培地
人工培地または合成培地の調製には、栄養素(有機・無機双方)、血清タンパク質、炭水化物、補因子、ビタミン、塩類に加え、O₂およびCO₂の気相成分の添加が含まれます[1]。
以下の機能のうち1つ以上を満たすために、さまざまな種類の人工培地が開発されてきた:1) 即時生存(正確なpHおよび浸透圧を有する平衡塩溶液)。2) 長期生存(さまざまな配合の有機化合物および/または血清を添加した平衡塩溶液)。 3) 無期限の発育。4) 特殊な機能。
人工培地には、以下の4つの明確な分類がある:
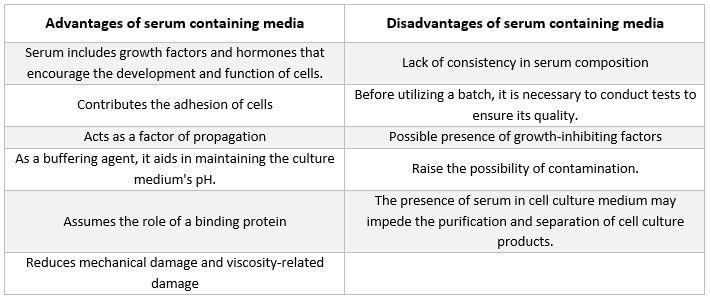
血清含有培地
動物細胞の培養に用いられる培地において、最も頻繁に見られる添加物はウシ胎児血清である。これは、可能な限り最良の増殖条件を実現するために、低コストの添加物として培養培地に添加される。 血清は、不安定または水に溶けない栄養素、ホルモンや成長因子、プロテアーゼ阻害剤、その他の物質の運搬体やキレート剤として機能するだけでなく、有害な分子を結合・中和する役割も果たします。
無血清培地
培地中に血清が含まれていることには多くの欠点があり、免疫学的研究において解釈上の重大な誤りを引き起こす可能性があります [2, 3]。これまでに、さまざまな無血清培地が開発されてきました [4, 5]。 これらの培地は一般的に、特定の細胞種の培養をサポートするように特別に調製されており、例えば、Thermo Fisher Scientific社の「Knockout Serum Replacement」や「Knockout DMEM」、Stem Cell Technologies社の幹細胞用「mTESR培地」などが挙げられる[6][7]。
さらに、これらの培地には、通常は血清によって供給される精製された成長因子、リポタンパク質、その他のタンパク質が、所定の量で配合されている[8]。これらの培地を構成する成分は十分に解明されているため、しばしば「定義済み培養培地」と呼ばれる。
化学的に定義された培地
これらの培地には、いかなる種類の汚染も受けていない超高純度の無機および有機成分が含まれています。また、成長因子などの純粋なタンパク質が添加されている場合もあります。
細菌や酵母の遺伝子改変に加え、特定の脂肪酸、ビタミン、コレステロール、アミノ酸を添加することで、その構成成分が生成される[9]。
無タンパク質培地
無タンパク質培地とは、タンパク質を一切含まず、代わりに非タンパク質成分のみを含む培地のことである。血清を添加した培地と比較して、タンパク質を添加しない培地を使用すると、細胞の増殖とタンパク質の発現が促進され、下流工程で生成される製品の精製が容易になる[10-12]。 MEMやRPMI-1640などの培地には、タンパク質は含まれていない。ただし、必要に応じてタンパク質サプリメントを添加する場合もある。
培養培地とその基本成分
市販の培養培地は粉末または液体の形で購入でき、多くの場合、アミノ酸、グルコース、塩類、ビタミン、その他の栄養補助成分など、様々な栄養素が含まれています。
これらの成分に対する必要量は細胞株ごとに異なり、こうした差異が培地の配合が数多く存在する理由となっている。各成分は特定の機能を担っており、それについては以下の段落で概説する:
緩衝系
最適な増殖条件を維持するためにはpHを制御する必要があり、これは通常、以下の2つの緩衝系のいずれかによって行われます:
天然の緩衝系
大気中のCO₂/H₂CO₃比は培液中のそれと同等であり、これが自然な緩衝メカニズムを生み出しています。この自然な緩衝メカニズムを維持するためには、培養物を5~10%のCO₂を含む空気環境下で保持する必要があり、これは多くの場合、CO₂インキュベーターを使用することで実現されます。 天然緩衝系を使用する最大の利点は、その安価さと安全性にあります。
HEPES
双性イオンであるHEPESを用いた化学的緩衝は、pH 7.2~7.4の範囲でより高い緩衝能力を持ち、ガス環境の制御を必要としません。特定の細胞種によっては、HEPESの投与量が多すぎると有害となる場合があります。 また、HEPES を含む培地は、蛍光灯による光毒性の影響を受けやすくなります [13]。
フェノールレッド
pH指示薬であるフェノールレッドは、市販の培養培地にしばしば含まれており、pHの継続的なモニタリングを可能にします。細胞が増殖するにつれて、細胞から生成される代謝産物がpHを変化させ、その結果、培地の色が変わります。 フェノールレッドは培地の色に二重の効果をもたらし、酸性pHでは黄色に、アルカリ性pHでは紫色に変化する。細胞培養の最適値であるpH 7.4では、培地は蛍光赤色に見える。
しかし、フェノールレッドにはいくつかの欠点がある。第一に、フェノールレッドは、主にエストロゲンをはじめとする多くのステロイドホルモンの作用を模倣する可能性がある[14]。そのため、乳腺組織のようなエストロゲン感受性細胞を研究する際には、フェノールレッドを含まない培地の使用が推奨される。 いくつかの無血清培地では、フェノールレッドの存在によりナトリウム・カリウムのバランスが乱される。培地に血清やウシ下垂体ホルモンを添加することで、この影響を打ち消すことができる[15]。第三に、フローサイトメトリー実験における検出が、フェノールレッドの存在によって妨げられる。
無機塩
ナトリウム、カリウム、カルシウムイオンなどの無機塩を含む培地は、浸透圧平衡の維持や膜電位の調節に寄与する。
アミノ酸
アミノ酸はタンパク質の基本構成要素であるため、これまでに考案されたあらゆる細胞培養培地の必須成分となっています。細胞は特定のアミノ酸を自ら生成することができないため、培養培地に必須アミノ酸を含めることが重要です。 これらは細胞の増殖に不可欠であり、その濃度によって達成可能な最大細胞密度が決まります。特に、必須アミノ酸であるL-グルタミンは極めて重要です。
L-グルタミンは、代謝のための二次的なエネルギー源として機能し、NAD、NADPH、およびヌクレオチドの生成に窒素を供給します。L-グルタミンは不安定なアミノ酸であり、時間の経過とともに細胞が利用できない形態に変化してしまうため、培地に添加する必要があります。
さらに、増殖過程で消費されたアミノ酸を補充するために、非必須アミノ酸を培地に添加することも可能です。増殖培地に非必須アミノ酸を添加することで、細胞の増殖が促進され、生存率も向上します。
炭水化物
糖類の形態をとる炭水化物は、主要なエネルギー源である。多くの培地には、一般的なグルコースやガラクトースに加え、マルトースやフルクトースも含まれている。
タンパク質およびペプチド
アルブミン、トランスフェリン、フィブロネクチンは、最も一般的に使用されるタンパク質およびペプチドである。これらは、血清を含まない培地において特に重要である。アルブミン、トランスフェリン、アプロチニン、フェトゥイン、フィブロネクチンなどは、タンパク質が豊富に含まれる血清中に見られるタンパク質の一部である。
アルブミンは血液中に存在する主要なタンパク質であり、その機能は、水、塩類、遊離脂肪酸、ホルモン、ビタミンなどの様々な物質を結合させ、異なる臓器や細胞間で輸送することにある。アルブミンには化学物質に結合する能力があるため、細胞培養培地から有害な化合物を除去するための有効な候補となる。
アプロチニンは、中性および酸性のpH条件下で安定しており、高温やタンパク質分解酵素による破壊にも耐性があるため、細胞培養システムにおける保護剤として用いられます。トリプシンをはじめとする多くのセリンプロテアーゼを阻害する能力を持っています。
フェトゥインは、成体の血清と比較して、胎児や新生動物の血清中でより高濃度に検出される可能性のある糖タンパク質である。さらに、セリンプロテアーゼ阻害剤としても作用する。タンパク質であるフィブロネクチンは、細胞接着の過程において不可欠な構成要素である。 トランスフェリンは、鉄を輸送し、細胞膜へ鉄を供給する役割を担うタンパク質である。
脂肪酸および脂質
これらは、血清が存在しない無血清培地において極めて重要な役割を果たす。
ビタミン
細胞の発達と増殖には、数多くのビタミンが必要です。ビタミンは細胞内で十分な量が産生できないため、組織培養においては栄養補助剤として不可欠です。
細胞培養において、血清はビタミンの主な供給源ですが、特定の細胞種に適した培地にするために、さまざまなビタミンが添加されることもあります。最も一般的なのは、増殖を促進するためにB群ビタミンが使用されることです。
微量元素
銅、亜鉛、セレン、トリカルボン酸中間体などの化学元素は、微量元素として知られています。微量元素は、血清に通常含まれている成分を補うために、血清を含まない培地に添加されることがよくあります。これらの元素は、健康な細胞の発達に不可欠な重要な化学成分です。 酵素活性など、多くの生化学反応は特定の微量栄養素に依存しています。
培地添加剤
特定の細胞株に推奨される完全培養液には、ベースライン培地や血清には含まれていない追加成分が必要です。これらの添加物は、細胞の増殖と適切な代謝機能をサポートします。
ホルモン、成長因子、シグナル伝達分子は特定の細胞株の適切な増殖に不可欠ですが、以下の注意事項を常に遵守する必要があります。添加物の添加により完全培養液の浸透圧が変化し、細胞の発育が阻害される可能性があるため、添加後は必ず浸透圧を確認することが推奨されます。 ほとんどの細胞株において、最適な浸透圧は260~320 mOSM/kgの範囲です。
抗生物質
抗生物質は、細菌や真菌による汚染の増殖を抑制するためにしばしば用いられますが [16]、細胞の増殖に必須というわけではありません。抗生物質は、マイコプラズマや耐性菌による汚染を隠してしまう可能性があるため、細胞培養における日常的な使用は推奨されません [17, 18]。
さらに、抗生物質は過敏な細胞の代謝を阻害する可能性がある。MilliporeSigma社およびLife Technologies社製のペニシリン・ストレプトマイシン配合剤が頻繁に使用される。 プラズモシンは、グリオーマ細胞株 TS603、TS516、および BT260 の培養に利用されており [19]、マイコプラズマ汚染の除去に有効であることが示されている (20)。
血清
血清には、アルブミン、成長因子、および増殖阻害因子がすべて含まれている。血清は、アミノ酸、タンパク質、ビタミン(特にA、D、E、Kなどの脂溶性ビタミン)、炭水化物、脂質、ホルモン、成長因子、ミネラル、微量元素を供給するため、細胞培養培地の最も重要な成分の一つである。
培養細胞の発育を促進するために、胎児牛由来および子牛由来の血清がしばしば利用される。胎児血清は成長因子を豊富に含んでおり、細胞のクローニングや感受性の高い細胞の培養に適している。一方、子牛血清は増殖促進能力が低いため、接触抑制実験に用いられる。 通常の培養液には、多くの場合2%から10%の血清が含まれています。培養液への血清添加には、以下の目的があります[21]:
-
血清は、細胞に必須の栄養素を供給する(溶液中およびタンパク質に結合した形態の両方)。
-
血清には、増殖促進や特殊な細胞活動に関与するいくつかの成長因子やホルモンが含まれている。
-
血清には、アルブミンやトランスフェリンなど、他の化学物質を細胞内に輸送する多くの結合タンパク質が含まれている。例えば、アルブミンは脂肪、ビタミン、ホルモンなどを細胞内に運ぶ。
-
また、フィブロネクチンなどのタンパク質も提供し、これらは基質への細胞接着を促進する。さらに、分裂前の細胞の拡大を助ける「スプレッディング要素」も生成する。
-
また、細胞内でのタンパク質分解を防ぐプロテアーゼ阻害剤も供給します。
-
また、Na+、K+、Zn2+、Fe2+などのミネラルも含まれています。
-
培地の粘度を高めるため、浮遊培養時の撹拌中に細胞が機械的損傷を受けるのを防ぎます。
-
また、緩衝剤としての役割も果たします。
参考文献
[1] Morgan J, Morton H, Parker R. 組織培養における動物細胞の栄養:合成培地に関する初期の研究。Proc Soc Exp Biol Med. 1950;73:1-8
[2] Kerbel R, Blakeslee D. 培養中の哺乳類細胞による胎児牛血清成分の急速な吸着。細胞特異的抗原に対する抗血清の研究における人工的要因の潜在的な原因。Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. 膝窩リンパ節試験を用いて検討した細胞媒介反応における人工的変化の潜在的な原因としての、細胞懸濁液調製用培地への血清添加. J Immunogenet. 1980;7:483-9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. 市販の無血清培地:ハイブリドーマの増殖とモノクローナル抗体の産生. J Immunol Methods. 1991;145:175-83
[5] Barnes D, Sato G. 無血清培地における培養細胞の増殖法. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O, et al. HSP70シャペロンは、RNAを含まないTDP-43を異方性を持つ核内液状球殻へと誘導する。Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, et al. ダウン症候群に起因する老化は、神経前駆細胞の核構造を破壊する。 Cell Stem Cell. 2022;29:116-130.e7
[8] Iscove N, Melchers F. リポ多糖に反応するBリンパ球の培養において、血清をアルブミン、トランスフェリン、および大豆脂質で完全に置換すること。J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. ハイブリドーマの増殖およびモノクローナル抗体産生のための、化学的に定義された無タンパク質培地の体系的な改良。J Biotechnol. 1996;45:111-23
[10] Darfler F. ハイブリドーマおよびその他の免疫系細胞の増殖のための無タンパク質培地. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. 血清フリー細胞培養:統一的なアプローチ. Cell. 1980;22:649-55
[12] Hamilton W, Ham R. 無タンパク質培地におけるチャイニーズハムスター細胞株のクローン増殖. In Vitro. 1977;13:537-47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. 光に曝露されたHEPES含有培養液の細胞毒性効果の解析. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. 組織培養培地中のフェノールレッドは弱いエストロゲンである:培養におけるエストロゲン応答性細胞の研究に関する示唆. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. 無血清培地の開発. In: Master JRW, 編. Animal Cell culture, 第3版. オックスフォード:オックスフォード大学出版局; 2000.
[16] Perlman D. 細胞培養培地における抗生物質の使用. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. 細胞培養におけるマイコプラズマ感染の拡大と制御. In Vitro. 1976;12:643-8
[18] Masters J, Stacey G. 培地の交換と細胞株の継代. Nat Protoc. 2007;2:2276-84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M, et al. ヒストン脱メチル化酵素KDM6Aは酸素を直接感知し、クロマチンおよび細胞の運命を制御する。 Science. 2019;363:1217-1222
[20] Molla Kazemiha V、Azari S、Amanzadeh A、Bonakdar S、Shojaei Moghadam M、 Habibi Anbouhi M 他. 細胞培養で一般的に使用される抗生物質と比較した、モリキュートに感染した様々な哺乳類細胞株に対する Plasmocin™ の有効性:現地での経験. Cytotechnology. 2011;63:609-20
[21] Kragh Hansen U. 血清アルブミンへのリガンド結合の分子的側面. Pharmacol Rev. 1981;33:17-53