
, L-alanyyli-L-glutamiiniliuos")
Tilavuus: 100 ml
Säilytys: ≤ –15 °C
Steriiliys: Sterilisoitu suodatus
Stable Glutamine Solution (L-Alanyl-L-Glutamine, 200 mM) on erittäin stabiili L-glutamiinin dipeptidimuoto, joka on suunniteltu korvaamaan perinteinen L-glutamiini soluviljelyalustoissa. L-glutamiini on välttämätön aminohappo ja tärkeä energianlähde viljellyille soluille, ja sillä on tärkeä rooli solujen kasvussa, aineenvaihdunnassa ja proteiinisynteesissä.
Käyttö ja edut
Tavallisissa nestemäisissä viljelyaineissa L-glutamiini hajoaa suhteellisen nopeasti 37 °C:ssa, mikä johtaa myrkyllisten sivutuotteiden, kuten ammoniumionien, muodostumiseen, jotka voivat vaikuttaa negatiivisesti solujen elinkelpoisuuteen ja kokeiden tuloksiin. Stabiili glutamiini ratkaisee tämän ongelman tarjoamalla hajoamattoman dipeptidimuodon, joka pysyy ennallaan viljelyolosuhteissa.
Solut pilkkovat dipeptidisidoksen entsymaattisesti vapauttaakseen L-glutamiinia tarpeen mukaan, mikä varmistaa jatkuvan tuoreen saannin ja estää haitallisten jätetuotteiden kertymisen. Tämä tekee ratkaisusta erityisen edullisen pitkäaikaisille soluviljelmille ja tiheille kasvujärjestelmille.
Koostumus ja käyttö
L-alanyyli-L-glutamiiniliuos valmistetaan soluviljelylaatuiseen veteen 200 mM:n pitoisuutena ja suodatetaan steriilisti kontaminaatiolle herkkiä sovelluksia varten. Se voidaan laimentaa suoraan täydelliseen elatusaineeseen kokeellisten vaatimusten mukaisesti. Säilytä ≤ –15 °C:ssa ja vältä toistuvia jäädyttämis-sulatusjaksoja tuotteen vakauden säilyttämiseksi.
Vain tutkimuskäyttöön. Ei saa käyttää diagnostisissa tai terapeuttisissa toimenpiteissä. Ei saa käyttää ihmisillä tai eläimillä.
")
Tilavuus: 100 ml
Säilytys: +2 °C – +8 °C
Steriiliys: Sterilisoitu suodatus
HEPES-puskuriliuos (1 M), joka tunnetaan myös nimellä N-2-hydroksietyylipiperatsiini-N-2-etaanisulfonihappo, on zwitterioninen orgaaninen puskuri, jota käytetään laajalti soluviljelyalustoissa. Se on suunniteltu ylläpitämään vakaita pH-olosuhteita fysiologisella alueella 6,7–8,6, mikä tukee optimaalista solutoimintaa in vitro -sovelluksissa.
Käyttö ja edut
HEPES tarjoaa luotettavan puskurointikyvyn soluviljelyjärjestelmissä, erityisesti kun soluja käsitellään CO₂-inkubaattorin ulkopuolella. 10–25 mM HEPES:n lisääminen viljelyalustaan parantaa pH-stabiilisuutta pitkien käsittelyjaksojen aikana ja auttaa ylläpitämään tasaisia koeolosuhteita.
Tämä puskuri on kalvonläpäisemätön, häiritsee biokemiallisia reaktioita minimaalisesti ja osoittaa vahvaa kemiallista ja entsymaattista stabiilisuutta. Nämä ominaisuudet tekevät siitä sopivan monenlaisiin soluviljely
- ja biokemiallisiin sovelluksiin.
Koostumus ja käyttö
Liuos toimitetaan 1 M:n konsentraattina, joka on valmistettu soluviljelylaatuista vedestä ja suodatettu steriiliksi käytettäväksi kontaminaatiolle herkissä ympäristöissä. Se voidaan laimentaa haluttuun käyttöpitoisuuteen sovelluksen vaatimusten mukaan. Säilytä +2 °C – +8 °C:ssa ja käsittele aseptisissa olosuhteissa tuotteen eheyden säilyttämiseksi.
Vain tutkimuskäyttöön. Ei saa käyttää diagnostisissa tai terapeuttisissa toimenpiteissä. Ei saa käyttää ihmisillä tai eläimillä.
, 250 g/l")
Tilavuus: 50 ml
Säilytys: +2 °C – +8 °C
Steriiliys: Sterilisoitu suodattamalla
D-(+)-glukoosiliuos (dekstroosiliuos) on steriili, käyttövalmis ravintolisä, joka sisältää luonnossa esiintyvää D-(+)-glukoosia, joka on solujen aineenvaihdunnan keskeinen komponentti. Glukoosi osallistuu tärkeisiin biologisiin prosesseihin, kuten energian tuotantoon, glykosylaatioon ja glykaanien muodostumiseen, jotka vaikuttavat solujen rakenteeseen ja toimintaan.
Käyttö ja edut
Tätä glukoosiliuosta käytetään laajalti lisäravinteena soluviljelyalustoissa ja lukuisissa solu
- ja molekyylibiologian sovelluksissa. Glukoosi on ensisijainen hiili
- ja energialähde, joka tukee solujen kasvua, lisääntymistä ja aineenvaihduntaa. Sen osallistuminen biosynteettisiin reaktioihin tekee siitä myös tärkeän normaalin solufysiologian ja kokeiden yhdenmukaisuuden ylläpitämisessä.
Koostumus ja käyttö
Liuos toimitetaan korkeana pitoisuutena, 250 g/l glukoosia, mikä mahdollistaa joustavan laimentamisen viljelyalustoihin kokeellisten vaatimusten mukaisesti. Se on steriilisuodatettu, jotta se sopii kontaminaatiolle herkkiin sovelluksiin. Säilytä +2 °C – +8 °C:ssa ja käsittele aseptisesti tuotteen laadun ja vakauden säilyttämiseksi.
Vain tutkimuskäyttöön. Ei saa käyttää diagnostisissa tai terapeuttisissa toimenpiteissä. Ei saa käyttää ihmisillä tai eläimillä.
, liuos 100x")
Tilavuus: 10 ml
Säilytys: +2 °C – +8 °C
Steriliteetti: Sterilisuodatettu
Insuliini-transferriini-seleeni (ITS) -liuos (100x) on kemiallisesti määritelty lisäaine, joka on suunniteltu monenlaisiin soluviljelykäyttöihin. Sitä käytetään yleisimmin lisäaineena perusviljelyalustoihin tukemaan solujen kasvua vähäseerumissa tai seerumittomissa olosuhteissa.
Käyttö ja edut
ITS-lisäaineemme tarjoaa välttämättömät komponentit, joita tarvitaan seerumittomien elatusaineiden optimaaliseen toimintaan. Lisäämällä ITS:ää tavanomaisiin ravintoaineisiin voidaan monien solulinjojen rutiinihoidossa tarvittavan sikiön naudan seerumin (FBS) määrää vähentää merkittävästi. Tämä auttaa minimoimaan seerumin käyttöön liittyvää vaihtelua ja ylläpitämään samalla tasaisen solujen kasvun ja elinkelpoisuuden.
Insuliini tukee solujen ravinteiden ottoa ja aineenvaihduntaa, transferriini helpottaa raudan kuljetusta ja seleeni edistää antioksidanttisuojausta ja entsymaattista aktiivisuutta. Yhdessä nämä komponentit edistävät tasapainoista solujen aineenvaihduntaa ja parantavat toistettavuutta määritellyissä viljelyjärjestelmissä.
Koostumus ja käyttö
Insuliini-transferriini-seleeni (ITS) toimitetaan 100-kertaisesti konsentroituna liuoksena Earle's Balanced Salt Solution (EBSS) -liuoksessa ilman fenolipunaviiniä. Tavallisissa käyttötarkoituksissa laimennetaan 1:100 suhteessa sopivaan peruskasvatusliuokseen suositellun käyttöpitoisuuden saavuttamiseksi. Säilytetään +2 °C – +8 °C:ssa ja käsitellään aseptisissa olosuhteissa tuotteen vakauden ja steriiliyden säilyttämiseksi.
Vain tutkimuskäyttöön. Ei saa käyttää diagnostisissa tai terapeuttisissa toimenpiteissä. Ei saa käyttää ihmisillä tai eläimillä.
")
Tilavuus: 5 ml
Säilytys: +2 °C – +8 °C
Steriliteetti: Sterilisuodatettu
Ihmisperäinen rekombinantti-insuliiniliuos on kemiallisesti määritelty lisäaine, jota käytetään yleisesti nisäkässolulinjojen, kuten kiinalaisen hamsterin munasarjasolujen (CHO-solujen), viljelyssä. Tämä soluviljelylaatuinen liuos sisältää Saccharomyces cerevisiae -hiivassa ilmentynyttä rekombinanttia ihmisperäistä insuliinia, mikä takaa korkean puhtauden ja tasaisen suorituskyvyn tutkimussovelluksissa.
Käyttö ja edut
Insuliinia lisätään rutiininomaisesti seerumittomiin ja kemiallisesti määriteltyihin elatusaineisiin solujen kasvun ja tuottavuuden edistämiseksi. Keskeisenä säätelyhormonina insuliini tukee glukoosin, aminohappojen ja rasvahappojen soluunottoa, hyödyntämistä ja varastointia. Se myös estää glykogeenin, proteiinien ja lipidien hajoamista, mikä parantaa solujen elinkelpoisuutta ja metabolista vakautta viljelyjärjestelmissä. Kemiallisesti määritelty koostumus tukee toistettavuutta ja minimoi vaihtelevuutta herkissä soluviljelyprosesseissa.
Biologiset ominaisuudet ja käyttö
Insuliini on kaksiketjuinen polypeptidihormoni, jota haiman saarekkeiden β-solut tuottavat luonnollisesti. Sen molekyylipaino on noin 5 800 Da. α
- ja β-ketjut ovat yhteydessä toisiinsa kahden ketjujen välisen disulfidisidoksen kautta, ja α-ketjussa on yksi ketjun sisäinen disulfidisidos. Soluviljelykäytössä liuosta tulee käsitellä aseptisissa olosuhteissa ja säilyttää +2 °C – +8 °C:ssa stabiilisuuden ja suorituskyvyn säilyttämiseksi.
Vain tutkimuskäyttöön. Ei saa käyttää diagnostisissa tai terapeuttisissa toimenpiteissä. Ei saa käyttää ihmisillä tai eläimillä.
")
Tilavuus: 100 ml
Säilytys: +2 °C – +8 °C
Steriiliys: Sterilisoitu suodatus
Natriumpyruvaattiliuos (100 mM) on steriili, käyttövalmis lisäaine, joka on suunniteltu tarjoamaan lisäenergialähde soluviljelyalustoille. Natriumpyruvaatti on tärkeä osa solujen energia-aineenvaihduntaa ja tukee metabolisesti aktiivisten ja nopeasti lisääntyvien solujen, kuten kasvainsolujen, kasvua. Lisäaine voi parantaa solujen elinkelpoisuutta ja auttaa ylläpitämään metabolista vakautta viljelyjärjestelmissä.
Käyttö ja edut
Tätä liuosta käytetään laajalti rutiininomaisessa soluviljelyssä rikastamaan elatusaineita pyruvaatilla ja edistämään optimaalisia kasvuolosuhteita. Se tukee ATP:n tuotantoa, voi auttaa vähentämään oksidatiivista stressiä ja parantaa viljeltyjen solujen aineenvaihduntaa. Tuote on valmistettu soluviljelylaadusta vedestä ja steriilisuodatettu, mikä takaa tasaisen laadun ja toistettavuuden tutkimustyössä.
Käyttö ja yhteensopivuus
Suositeltu lopullinen pitoisuus useimmille soluviljelykäytöille on 1 mM natriumpyruvaatti, joka saavutetaan laimentamalla 100 mM:n kantaliuos 1:100 täydelliseen viljelyväliaineeseen. Liuos on yhteensopiva monien perusväliaineiden ja nisäkässolulinjojen kanssa. Säilytä +2 °C – +8 °C:ssa ja suojaa kontaminaatiolta tuotteen vakauden säilyttämiseksi.
Vain tutkimuskäyttöön. Ei saa käyttää diagnostisissa tai terapeuttisissa toimenpiteissä. Ei saa käyttää ihmisillä tai eläimillä.
")
Tilavuus: 100 ml Säilytys: ≤-15°C Steriliteetti: Steriili suodatettu
Antibiootti-/antimykoottiliuos (100x) on steriili, käyttövalmis konsentraatti, joka on suunniteltu vähentämään mikrobikontaminaatioriskiä soluviljelyssä ja siihen liittyvissä laboratoriosovelluksissa. Tämä 100x-liuos sisältää vakiintuneen yhdistelmän penisilliiniä, streptomysiiniä ja amfoterisiini B:tä, joka tarjoaa laajakirjoisen antimikrobisen aktiivisuuden grampositiivisia ja gramnegatiivisia bakteereja, hiivoja ja säikeisiä sieniä vastaan. Valmiste soveltuu käytettäväksi eukaryoottisissa soluviljelmissä, bakteerialustoissa ja muissa kontaminaatioherkissä järjestelmissä, mikä tukee puhtaita ja johdonmukaisia laboratoriotoimintoja.
Käyttö ja hyödyt Rutiininomaisiin tutkimusprotokolliin optimoitua liuosta käytetään laajalti aseptisten olosuhteiden ylläpitämiseen soluviljelytyöprosesseissa. Se tarjoaa luotettavan suorituskyvyn kontaminaatioherkissä ympäristöissä ja auttaa tutkijoita vähentämään mikrobien liikakasvun riskiä vaarantamatta solujen terveyttä tai kokeiden toistettavuutta. Steriilisuodatettu koostumus poistaa tarpeen ylimääräisille liuotusvaiheille, mikä tukee virtaviivaista väliaineen valmistusta ja vähentää vaihtelua päivittäisissä laboratoriomenetelmissä.
Käyttö ja yhteensopivuus Saavuttaaksesi vakiotyöpitoisuudet laimenna liuos 1:100 täydelliseen kasvatusalustaan. Tuote on yhteensopiva monenlaisten nisäkässolulinjojen ja perusmedioiden kanssa. Jatkuvan varastosaatavuuden ansiosta tutkijat hyötyvät toimitusten luotettavasta jatkuvuudesta ja logistiikkasuunnittelun yksinkertaistamisesta. Liuos on säilytettävä ≤ -15 °C:ssa ja suojattava toistuvilta jäädytys-sulatussykleiltä stabiilisuuden säilyttämiseksi. Vain tutkimuskäyttöön. Ei käytettäväksi diagnostisissa tai terapeuttisissa toimenpiteissä. Ei käytettäväksi ihmisillä tai eläimillä.
")
Tilavuus: 100 ml Säilytys: Säilytä: säilytys: +2°C
- +8°C Steriili suodatettu
MEM Non-Essential Amino Acids (100x) on steriili lisäravinne, joka on suunniteltu parantamaan solujen kasvua ja elinkelpoisuutta nisäkässolujen soluviljelyjärjestelmissä. Koostumus vastaa 100-kertaista konsentraattia ei-essentiaalisista aminohapoista, joita esiintyy standardissa MEM:ssä (Minimum Essential Medium), mikä mahdollistaa perusväliaineen suoran täydentämisen minimaalisella valmistelutyöllä.
Sovellus ja hyödyt Tämä lisäaine tarjoaa ylimääräisen aminohappopoolin nopeasti lisääntyville soluille tai solulinjoille, jotka ovat menettäneet kykynsä syntetisoida de novo muita kuin välttämättömiä aminohappoja. Keventämällä biosynteesin aiheuttamaa metabolista taakkaa se tukee parempaa kasvukinetiikkaa, pidempää elinkelpoisuutta ja suurempaa kokeellista johdonmukaisuutta
- erityisesti ravinteiden suhteen herkissä tai suuren tiheyden viljelmissä.
Koostumus ja käyttö Liuos sisältää glysiiniä, L-alaniinia, L-asparagiinia, L-asparagiinihappoa, L-glutamiinihappoa, L-proliinia ja L-seriiniä. Se on yhteensopiva MEM:n ja useimpien muiden vakiomedioiden kanssa. Käyttöä varten laimennetaan 1:100 lopulliseen kasvatusmediaan. Tämä tuote on steriilisti suodatettu ja käyttövalmis ilman ylimääräisiä käsittelyvaiheita. Vain tutkimuskäyttöön. Ei käytettäväksi diagnostisissa tai terapeuttisissa toimenpiteissä. Ei käytettäväksi ihmisillä tai eläimillä.

Accutase on käyttövalmis, steriilisti suodatettu solujen irrotusliuos, joka on suunniteltu hellävaraiseksi vaihtoehdoksi trypsiinille/EDTA:lle kiinnittyneiden nisäkässolujen irrottamiseksi tavallisista kudosviljelymuovivälineistä ja tarttuviksi pinnoitetuista pinnoista. Se yhdistää proteolyyttisen ja kollageenolyyttisen entsyymiaktiivisuuden tasapainotetussa suolaliuoksessa tehokkaan mutta hallitun irrottamisen aikaansaamiseksi, säilyttäen solupinnan proteiinit ja tukien korkeaa elinkelpoisuutta siirron jälkeen sekä nopeaa uudelleenkiinnittymistä.
Accutase-valmiste perustuu Dulbeccon fosfaattipuskuroituun suolaliuokseen (DPBS), jossa on EDTA:ta ja fenolipunainen visuaalisena pH-indikaattorina. Entsyymit ovat peräisin muista kuin nisäkkäistä tai bakteereista, minkä vuoksi Accutase sopii erityisen hyvin kantasolututkimukseen, rokotetyönkulkuihin ja muihin sovelluksiin, joissa eläin
- tai mikrobiperäisten kontaminanttien määrä on minimoitava. Liuos estää itsensä 37 °C:ssa, joten irrottamisen jälkeen ei tarvita neutraloivia reagensseja tai seerumia sisältävää elatusainetta – solut voidaan siirtää suoraan tuoreeseen elatusaineeseen.
Tärkeimmät ominaisuudet
Käyttövalmis 1x steriilisuodatettu neste – laimentamista tai rekonstituointia ei tarvita
Yhdistetty proteolyyttinen ja kollageenolyyttinen entsyymiaktiivisuus hellävaraista irrottamista varten
Jokainen erä on standardoitu määriteltyyn irrottamisaktiivisuuteen eräkohtaisen yhdenmukaisuuden varmistamiseksi
Entsyymit eivät ole peräisin nisäkkäistä tai bakteereista
Inhiboi itsensä 37 °C:ssa – neutralointiliuosta ei tarvita
Valmistettu Dulbeccon PBS:ssä EDTA:n kanssa
Sisältää fenolipunaisen visuaalisena pH-indikaattorina
pH 6,8–7,8
Tyypillisiä käyttökohteita
Accutase irrottaa hellävaraisesti monenlaisia kiinnittyneitä ja herkkiä solutyyppejä, mukaan lukien ihmisen alkion kantasolut (hESCs), ihmisen indusoidut pluripotentit kantasolut (iPSCs), hermoston kantasolut, primaariset neuronit sekä rutiinikäytössä olevat kiinnittyneet solulinjat, kuten HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 ja A549. Tyypillisiä käyttötapauksia ovat:
Adheesiivisten nisäkässolujen rutiininen aliviljely ja siirto
HES-solujen, iPS-solujen ja muiden herkkien solulinjojen hellävarainen yksittäisten solujen irrottaminen
Näytteiden valmistelu virtaussytometriaa ja FACS-analyysiä varten
Solupintamarkkerien analysointi, jossa epitopin eheys on tärkeää
Solujen migraatio-, proliferaatio
- ja apoptoosimääritykset
Lepotilan määritykset seerumin puutteella ja onkogeenien transfektiotutkimukset
Kasvainsolujen ja hermostopäällysteen solujen migraatiomääritykset
Tuotannon laajentaminen bioreaktoriprosesseissa
Rutiinityössä levitä noin 10 ml Accutasea 75 cm2:n viljelypinta-alaa kohti ja inkuboi 5–10 minuuttia huoneenlämmössä. Optimaalinen inkubointiaika on määritettävä kullekin solulinjalle erikseen, eikä se saa ylittää yhtä tuntia. Ennen lisäystä huuhtele solukerros Ca2+/Mg2+-vapaalla suolaliuoksella, kuten kalsiumia ja magnesiumia sisältämättömällä DPBS:llä, jotta poistat jäljellä olevan seerumin ja kaksiarvoiset kationit.
Käsittely ja säilytys
Säilytä avaamatonta pulloa pakastettuna -15 °C:ssa tai sitä alemmassa lämpötilassa. Sulata joko huoneenlämmössä tai yön yli +2 °C:ssa – +8 °C:ssa. Älä sulata Accutasea 37 °C:n vesihauteessa, sillä korkeat lämpötilat heikentävät entsyymiaktiivisuutta. Sulatuksen jälkeen liuosta voidaan säilyttää enintään 2 kuukautta +2 °C:ssa – +8 °C:ssa; älä säilytä huoneenlämmössä. Älä lämmitä reagenssia 37 °C:seen ennen käyttöä – lisää se suoraan pestyihin soluihin huoneenlämmössä. Pitkän säilyvyyden takaamiseksi suositellaan kertakäyttöisten alikvojen käyttöä, jotta vältytään toistuvilta sulatusjaksoilta. Työskentele aina aseptisissa olosuhteissa.
Laatu
Valmistettu tiukkojen laatustandardien mukaisesti. Jokainen Accutase-erä on steriilisuodatettu ja testattu steriilisyyden, pH:n, ulkonäön ja dissosiaatioaktiivisuuden osalta, jotta varmistetaan erästä toiseen yhdenmukainen ja toistettava suorituskyky.
Tuotetiedot
Tekniset tiedot
Yksityiskohdat
TuotetyyppiSolujen irrottamis-/dissosiaatioreagenssi
MuotoSteriilisuodatettu neste, käyttövalmis
Tilavuus100 ml
Käyttökonsentraatio1x (käyttövalmis)
EntsyymiaktiivisuusYhdistetty proteolyyttinen ja kollageenolyyttinen
Entsyymin alkuperäEi-nisäkäs
- ja ei-bakteeriperäinen
PuskurijärjestelmäDulbeccon PBS ja EDTA
pH-indikaattoriFenolipunainen
pH-alue6,8 – 7,8
UlkonäköKirkas, vaaleanpunainen tai oranssi liuos
Säilytyslämpötila-15 °C tai alle
Stabiilius sulatuksen jälkeenEnintään 2 kuukautta +2 °C – +8 °C:ssa
Suositeltu käyttömäärä~10 ml / 75 cm² viljelypintaa
Tyypillinen inkubointiaika5–10 minuuttia huoneenlämmössä
KuljetusolosuhteetPakastettu kuivajäällä
KäyttötarkoitusVain tutkimuskäyttöön ja jatkovalmistukseen
Koostumus (koostumus litraa kohti)
Komponentti
Pitoisuus (mg/l)
Epäorgaaniset suolat
Natriumkloridi (NaCl)8000,00
Dinatriumvetyfosfaatti (Na2HPO4)1150,00
Kaliumkloridi (KCl)200,00
Kaliumdihydrogeenifosfaatti (KH2PO4)200,00
Muut komponentit
EDTA · 4Na (tetranatrium-EDTA)220,00
Fenolipunainen3,00
Patentoitu entsyymiseos (proteolyyttinen ja kollageenolyyttinen aktiivisuus)1x
Accutase on Innovative Cell Technologies, Inc:n rekisteröity tavaramerkki.
, w: 2 mM L-Glutamiinia, w: 2,2 g/L NaHCO3, w: EBSS")
Tämä käyttövalmis, steriilisti suodatettu nestemäinen formulaatio on täydennetty Earle’s Balanced Salt Solution (EBSS) -liuoksella, 2 mM L-glutamiinilla, D-glukoosilla (1,0 g/l) ja 2,2 g/l natriumbikarbonaatilla (NaHCO3), minkä ansiosta se sopii käytettäväksi CO2-säädellyssä inkubaattoriympäristössä (tyypillisesti 5 % CO2). Mukana oleva fenolipunainen toimii pH-indikaattorina, mikä mahdollistaa väliaineen tilan kätevän visuaalisen seurannan soluviljelyn aikana.
Tärkeimmät ominaisuudet
Klassinen Eagle’s MEM -koostumus, jossa on Earle’s Balanced Salt Solution (EBSS)
Sisältää 2 mM L-glutamiinia – valmis välittömään käyttöön
2,2 g/l natriumbikarbonaattia – puskuroitu 5 %:n CO2-inkubaatiota varten
Pääasiallisena hiilen lähteenä D-glukoosi (1,0 g/l)
Fenolipunainen pH-indikaattorina
Ilman HEPES:ää ja ilman natriumpyruvaattia
Steriilisuodatettu nestemäinen elatusaine, käyttövalmis
pH 7,0 – 7,6
Tyypillisiä sovelluksia
EMEM tukee monenlaisten nisäkässolulinjojen viljelyä, mukaan lukien HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 ja monet primaarisolut. Yleisiä käyttökohteita ovat:
Adherenttien solulinjojen rutiinihoito ja laajentaminen
Virusten lisääminen ja rokotetuotannon työnkulut
Sytotoksisuus
- ja bioassay-sovellukset
Transfektio
- ja proteiini-ilmentymistutkimukset
Solubiologian ja molekyylibiologian perustutkimus
Optimaalisen solujen kasvun saavuttamiseksi EMEM-alustaan lisätään tyypillisesti 5–10 % naudan sikiöseerumia (FBS) ja, solulinjasta riippuen, ei-välttämättömiä aminohappoja (NEAA) sekä antibiootteja, kuten penisilliiniä/streptomysiiniä.
Käsittely ja säilytys
Säilytä avaamaton pullo +2 °C – +8 °C:ssa, valolta suojattuna. Avaamisen jälkeen käytä aseptisissa olosuhteissa. L-glutamiini liuoksessa hajoaa vähitellen – suosittelemme käyttämään väliainetta 4 viikon kuluessa avaamisesta parhaan suorituskyvyn saavuttamiseksi tai lisäämään tuoretta L-glutamiinia ennen käyttöä, jos sitä säilytetään pidempään. Anna väliaineen lämmetä 37 °C:seen ennen sen lisäämistä soluihin.
Laatu
Valmistettu tiukkojen laatustandardien mukaisesti. Jokainen erä testataan steriilisyyden, pH:n, osmolaliteetin ja endotoksiinipitoisuuden osalta, jotta varmistetaan tasainen suorituskyky soluviljelykäytöissä.
Tuotetiedot
Tekniset tiedot
Yksityiskohdat
TuotetyyppiMEM
TuoteryhmäSoluviljelyalustat
MuotoNestemäinen
SteriiliKyllä
Koko500 ml
L-glutamiiniSisältää L-glutamiinia (2 mM)
GlukoosiSisältää glukoosia (1,0 g/l)
NatriumbikarbonaattiNaHCO3:lla (2,2 g/l)
HEPESIlman HEPES:ää
Natrium-pyruvaattiIlman natriumpyruvaattia
FenolipunainenFenolipunalla
SuolaliuosEarlen tasapainotettu suolaliuos (EBSS)
pH7,0 – 7,6
EndotoksiinipitoisuusEi määritelty
Säilytys+2 °C – +8 °C
Koostumus (litraa kohti)
Ainesosa
Pitoisuus (mg/l)
Epäorgaaniset suolat
Kalsiumkloridi · 2H2O265,00
Magnesiumsulfaatti97,72
Kaliumkloridi400,00
Natriumkloridi6 800,00
Natriumdihydrogeenifosfaatti, vedetön122,00
Natriumbikarbonaatti (NaHCO3)2 200,00
Aminohapot
L-arginiini · HCl126,00
L-kystiini · 2HCl31,30
L-glutamiini292,00
L-histidiini · HCl · H2O42,00
L-isoleusiini52,00
L-leusiini52,00
L-lysiini · HCl72,50
L-metioniini15,00
L-fenyylialaniini32,00
L-treoniini48,00
L-tryptofaani10,00
L-tyrosiini · 2Na · 2H2O51,90
L-valiini46,00
Vitamiinit
D-kalsiumpantotenaatti1,00
Koliinikloridi1,00
Foolihappo1,00
myo-inositoli2,00
Nikotiiniamidi1,00
Pyridoksaali · HCl1,00
Riboflaviini0,10
Tiamiini · HCl1,00
Muut komponentit
D(+)-glukoosi1 000,00
Fenolipunainen10,00

- ja eläinsolut, mikä tekee siitä välttämättömän työkalun erilaisiin tutkimussovelluksiin. Freeze Medium CM-1 on koostettu huolellisesti tasapainotetusta yhdistelmästä kryosuojakemikaaleja ja välttämättömiä ravintoaineita, ja se minimoi jääkiteiden muodostumisen ja solustressin jäädytysprosessin aikana, jolloin solujen eheys säilyy.
Freeze Medium CM-1:n tärkeimpiä ominaisuuksia ovat mm:
Laaja yhteensopivuus: Tehokas monenlaisille solutyypeille, mukaan lukien primaarisolut, kantasolut ja vakiintuneet solulinjat.
Korkea elinkelpoisuus: Optimoitu maksimoimaan solujen palautuminen ja elinkelpoisuus sulatuksen jälkeen, mikä takaa luotettavat koetulokset.
Käyttövalmis: Kätevästi valmisteltu ja steriloitu välitöntä käyttöä varten, mikä vähentää valmisteluaikaa ja kontaminaatioriskiä.
Parannettu stabiilisuus: Säilyttää tasaisen suorituskyvyn tavanomaisissa kryosäilytysolosuhteissa, mikä takaa toistettavat tulokset.
Pitkä säilyvyysaika: CM-1 on seerumia sisältävä, käyttövalmis kryosäilytysmedia, jota voidaan säilyttää jääkaapissa jopa vuoden ajan.
CM-1:n käyttö solujen pakastamiseen
Jos haluat käyttää CM-1:tä sekä adheesiosolujen että suspensiosolujen jäädyttämiseen, toimi seuraavasti
Adheesiosolut pestään ja irrotetaan viljelyalustasta. Jos kyseessä ovat suspensiosolut, siirry suoraan seuraavaan vaiheeseen.
Laske solut varmistaaksesi, että niiden pitoisuus on oikea.
Sentrifugoi solut pelletiksi ja suspendoi ne uudelleen CM-1-pakastusmediumiin.
Siirretään resuspendoidut solut kryopulloihin.
Käytä hidasta pakastusmenetelmää ennen solujen siirtämistä pitkäaikaissäilytykseen
Menetelmä
Kuvaus
Vaiheet
❄️
Manuaalinen jäädytys
Vaiheittainen menetelmä, jossa lämpötilaa alennetaan asteittain solujen elinkelpoisuuden varmistamiseksi
1️⃣ Aseta solut pakastusmediumiin 4 °C:n pakastimeen 40 minuutiksi.
2️⃣ Siirretään -80 °C:n pakastimeen 24 tunniksi.
3️⃣ Säilytä solut nestemäisessä typessä pitkäaikaissäilytystä varten
❄️
Mr. Frostyn käyttäminen
Kätevä laite, joka mahdollistaa hallitun pakastusnopeuden ilman sähkövirtaa
1️⃣ Valmistele solut kryopulloihin pakastusmediumilla.
2️⃣ Aseta kryopullot Mr. Frosty -astiaan.
3️⃣ Säilytä -80 °C:ssa 24 tuntia ennen siirtämistä nestemäiseen typpeen
❄️
Pakastin, jonka nopeus on säädetty
Thermo Fisherin tai muiden valmistajien korkean tarkkuuden pakastin, joka on suunniteltu lämpötilan hallittuun alentamiseen
1️⃣ Ohjelmoi laite alentamaan lämpötilaa asteittain.
2️⃣ Aseta valmistetut solut pakastimeen.
3️⃣ Siirrä solut pakastussyklin jälkeen nestemäiseen typpeen
Säilytä kryopullot alle -130 °C:n lämpötilassa tai nestemäisessä typessä pitkäaikaissäilytystä varten.
Ainesosat
Sisältää FBS:ää, DMSO:ta, glukoosia, suoloja
Puskurikyky: pH = 7,2-7,6
Cytionin Freeze Medium CM-1 tarjoaa luotettavan ratkaisun kryosäilytykseen, joka takaa solujen korkean elinkelpoisuuden ja toimivuuden sulatuksen jälkeen monenlaisissa tutkimussovelluksissa.

Ham's F-12K (Kaighn's) Medium on huolellisesti muotoiltu soluviljelyolosuhteiden optimoimiseksi. Sen koostumus on rikastettu, ja se sisältää runsaasti välttämättömiä komponentteja, kuten aminohappoja ja natriumpyruvaattia, sekä lisäaineita, kuten putressiiniä, tymidiiniä, hypoksantiiniä ja sinkkiä. Näiden lisäysten avulla tutkijat voivat täydentää väliaineen minimaalisella seerumilla tai tietyille solutyypeille määritellyillä komponenteilla, mikä helpottaa täsmällisten koeolosuhteiden luomista.
Ham's F-12K (Kaighn's) Medium ei sisällä proteiineja eikä kasvutekijöitä. Näin ollen kasvutekijöillä ja naudan sikiöseerumilla (FBS) täydentäminen on usein tarpeen, jolloin tutkijat voivat räätälöidä väliaineen tiettyjen solulinjojensa vaatimusten mukaiseksi. Optimaalisen suorituskyvyn saavuttamiseksi FBS:n pitoisuus on optimoitava huolellisesti kullekin solulinjalle, jotta varmistetaan optimaalinen kasvu ja toimivuus.
Fysiologisen pH:n ylläpitämiseksi Ham's F-12K (Kaighn's) -mediumissa käytetään natriumbikarbonaattipuskurijärjestelmää (2,5 g/l), mikä edellyttää kontrolloitua 5-10 %:n hiilidioksidipitoisuutta viljelyn aikana. Näin varmistetaan, että väliaineen pH pysyy solujen kasvun ja elinkelpoisuuden kannalta ihanteellisella alueella
Laadunvalvonta
pH = 7,2 +/
- 0,02 20-25 °C:ssa.
Jokainen erä on testattu steriiliyden sekä mykoplasman ja bakteerien puuttumisen varalta.
Ylläpito
Säilytetään jääkaapissa +2°C
- +8°C:ssa pimeässä. Pakastaminen sekä lämmittäminen +37°C:seen asti vähentävät tuotteen laatua.
Älä lämmitä elatusainetta yli 37°C:een tai käytä hallitsemattomia lämmönlähteitä (esim. mikroaaltolaitteita).
Jos vain osa väliaineesta halutaan käyttää, poista tämä määrä pullosta ja lämmitä se huoneenlämmössä.
Kaikkien väliaineiden paitsi perusmediumin säilyvyysaika on 8 viikkoa valmistuspäivästä.
Koostumus
Ainesosat
mg/l
Epäorgaaniset suolat
Kalsiumkloridi x 2H2O
135,24
Kupari(II)sulfaatti x 5H2O
0,00
Rauta(II)sulfaatti x 7H2O
0,83
Magnesiumkloridi x 6H2O
105,72
Magnesiumsulfaatti x 7H2O
394,49
Kaliumkloridi
283,29
Kaliumdivetyfosfaatti
58,52
Natriumkloridi
7597,20
dinatriumvetyfosfaattivedetön
115,02
Sinkkisulfaatti x 7H2O
0,14
Muut aineosat
D(+)-glukoosi vedetön
1260,00
Hypoksantiini
4,08
DL-α-Lipoiinihappo
0,21
Fenolipunainen
3,00
Putressiini x 2HCl
0,32
Natriumpyruvaatti
220,00
NaHCO3
2500,00
Tymidiini
0,73
Aminohapot
L-alaniini
17,82
L-arginiini x HCl
421,40
L-asparagiini x H2O
30,02
L-asparagiinihappo
26,62
L-kysteiini x HCl x H2O
70,24
L-Glutamiini
292,20
L-glutamiinihappo
29,42
Glysiini
15,01
L-histidiini x HCl x H2O
41,92
L-isoleusiini
7,87
L-leusiini
26,24
L-lysiini x HCl
73,04
L-metioniini
8,95
L-Fenyylialaniini
9,91
L-proliini
69,06
L-Seriini
21,02
L-treoniini
23,82
L-tryptofaani
4,08
L-tyrosiini
10,87
L-valiini
23,42
Vitamiinit
D(+)-Biotiini
0,07
D-kalsiumpantotenaatti
0,48
Koliinikloridi
13,96
Foolihappo
1,32
myo-Inositoli
18,02
Nikotiiniamidi
0,04
Pyridoksiini x HCl
0,06
Riboflaviini
0,04
Tiamiini x HCl
0,34
B12-vitamiini
1,36

Fosfaattipuskuroitu suolaliuos (PBS) on laajalti käytetty puskuriliuos biologisessa ja kemiallisessa tutkimuksessa. Sillä on ratkaiseva merkitys pH-tasapainon ja osmolaarisuuden ylläpitämisessä erilaisten kokeellisten toimenpiteiden aikana, mukaan lukien kudosten käsittely ja soluviljely. PBS-liuoksemme on valmistettu huolellisesti erittäin puhtaista ainesosista vakauden ja luotettavuuden varmistamiseksi jokaisessa kokeessa. PBS-liuoksemme osmolaarisuus ja ionipitoisuudet jäljittelevät läheisesti ihmiskehon osmolaarisuutta ja ionipitoisuuksia, joten se on isotoninen ja myrkytön useimmille soluille.
PBS-liuoksemme koostumus
PBS-liuoksemme on pH-säädetty sekoitus erittäin puhdasta laatua olevia fosfaattipuskureita ja suolaliuoksia. Se sisältää 1-kertaisena käyttökonsentraationa:
8000 mg/l natriumkloridia (NaCl)
200 mg/l kaliumkloridia (KCl)
1150 mg/L natriumfosfaattidibasinen vedetön (Na2HPO4)
200 mg/L Kaliumfosfaatti, monobasinen, vedetön (KH2PO4)
Tämä koostumus takaa optimaalisen pH
- ja ionitasapainon, joka soveltuu monenlaisiin biologisiin sovelluksiin.
PBS-liuoksemme käyttökohteet
PBS-liuoksemme soveltuu erinomaisesti erilaisiin biologisen tutkimuksen sovelluksiin. Sen isotonisten ja myrkyttömien ominaisuuksien ansiosta se soveltuu aineiden laimentamiseen ja solusäiliöiden huuhteluun. EDTA:ta sisältävät PBS-liuokset irrottavat tehokkaasti kiinnittyneet ja kyhmyiset solut. PBS:ään ei kuitenkaan pitäisi lisätä kaksiarvoisia metalleja, kuten sinkkiä, sillä ne voivat aiheuttaa saostumista. Tällaisissa tapauksissa suositellaan Goodin puskureita. Lisäksi PBS-liuoksemme on hyväksyttävä vaihtoehto virusten kuljetusalustalle RNA-virusten, kuten SARS-CoV-2:n, kuljetuksessa ja säilytyksessä.
Laadunvalvonta
Steriilisuodatettu
Varastointi ja säilyvyys
Säilytetään +2 °C:n ja +25 °C:n välillä valolta suojattuna.
Kun se on avattu, säilytä 2°C-25°C:ssa ja käytä 24 kuukauden kuluessa.
Kuljetusolosuhteet
Ympäristön lämpötila
Huolto
Säilytettävä jääkaapissa +2°C
- +8°C:ssa pimeässä. Vältä pakastamista ja usein tapahtuvaa lämmittämistä +37°C:een, koska se heikentää tuotteen laatua.
Älä lämmitä väliaineen lämpötilaa yli 37°C äläkä käytä kontrolloimattomia lämmönlähteitä, kuten mikroaaltolaitteita.
Jos väliaineesta käytetään vain osa, poista tarvittava määrä ja lämmitä se huoneenlämpöiseksi ennen käyttöä.
Koostumus
Luokka
Ainesosat
Pitoisuus (mg/l)
Suolat
Kaliumkloridi
200
Kaliumfosfaattimonobasinen vedetön kaliumfosfaatti
200
Natriumkloridi
8000
Vedetön dibasinen natriumfosfaatti
1150

Alun perin se oli suunniteltu tukemaan ihmisen leukemiasolujen kasvua sekä suspensio
- että monokerrosviljelmissä, mutta tutkijoiden ja kaupallisten toimittajien tekemien muutosten myötä RPMI 1640 -medium on kehittynyt sopivaksi monille erilaisille nisäkässoluille. Se on poikkeuksellisen yhteensopiva sellaisten solulinjojen kanssa kuin HeLa, Jurkat, MCF-7, PC12, PBMC, astrosyytit ja karsinoomat.
RPMI 1640 Medium erottuu muista soluviljelymedioista ainutlaatuisen koostumuksensa ansiosta. Se sisältää huomattavan määrän fosfaattia, aminohappoja ja vitamiineja. Erityisesti se sisältää biotiinia, B12-vitamiinia ja PABA:ta, joita ei ole Eagle's Minimal Essential Mediumissa tai Dulbecco's Modified Eagle Mediumissa. Lisäksi RPMI 1640 -mediassa on huomattavasti korkeammat inositoli
- ja koliinivitamiinien pitoisuudet. Se ei kuitenkaan sisällä proteiineja, lipidejä tai kasvutekijöitä. Näin ollen solujen kasvulle optimaalisten olosuhteiden luomiseksi tarvitaan yleensä 10 %:lla naudan sikiöseerumia (FBS).
RPMI 1640 -mediumin puskurointijärjestelmä perustuu natriumbikarbonaattiin, ja fysiologisesti sopivan pH:n ylläpitäminen edellyttää 5-10 %:n CO2-ympäristöä. Pelkistävän aineen, glutationin, sisältyminen tähän väliaineeseen erottaa tämän väliaineen edelleen muista.
Laadunvalvonta
Steriilisuodatettu
Varastointi ja säilyvyys
Säilytetään +2°C
- +8°C:ssa valolta suojattuna.
Kun se on avattu, säilytä 4 °C:ssa ja käytä 6-8 viikon kuluessa.
Toimitusolosuhteet
Ympäristön lämpötila
Huolto
Säilytettävä jääkaapissa +2°C
- +8°C:ssa pimeässä. Vältä pakastamista ja usein tapahtuvaa lämmittämistä +37°C:seen, koska se heikentää tuotteen laatua.
Älä lämmitä väliaineen lämpötilaa yli 37°C äläkä käytä kontrolloimattomia lämmönlähteitä, kuten mikroaaltolaitteita.
Jos väliaineesta käytetään vain osa, poista tarvittava määrä ja lämmitä se huoneenlämpöiseksi ennen käyttöä.
Koostumus
Luokka
Ainesosat
Pitoisuus (mg/l)
Aminohapot
Glysiini
10.00
L-Alanyyli-L-Glutamiini
434.40
L-arginiini
200.00
L-asparagiiniH2O
56.82
L-asparagiinihappo
20.00
L-kystiini 2HCl
65.20
L-glutamiinihappo
20.00
L-histidiini HClH2O
20.27
L-hydroksi-L-proliini
20.00
L-isoleusiini
50.00
L-leusiini
50.00
L-lysiini HCl
40.00
L-metioniini
15.00
L-Fenyylialaniini
15.00
L-Proliini
20.00
L-Seriini
30.00
L-treoniini
20.00
L-tryptofaani
5.00
L-tyrosiini 2Na 2H2O
28.83
L-valiini
20.00
Vitamiinit
p-Aminobentsoehappo
1.00
D-Biotiini
0.20
Koliinikloridi
3.00
D-kalsium-pantotenaatti
0.25
Foolihappo
1.00
myo-Inositoli
35.00
Nikotiiniamidi
1.00
Pyridoksiini HCl
1.00
Riboflaviini
0.20
Tiamiini HCl
1.00
B12-vitamiini
0.005
Epäorgaaniset suolat
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Muut komponentit
D-glukoosi
2000.00
L-Glutationi pelkistetty
1.00
Fenolipunainen natriumsuola
5.30
, w: 3,1 g/L glukoosia, w: 2,5 mM L-glutamiinia, w: 15 mM HEPES, w: 0,5 mM natriumpyruvaattia, w: 1,2 g/L NaHCO3")
Tämä ainutlaatuinen koostumus yhdistää Dulbeccon modifioidun Eagle-alustan (DMEM) ja Ham's F-12:n (Ham's Nutrient Mixture F-12) tarkassa 1:1-suhteessa. L-glutamiinin lisääminen parantaa sen koostumusta entisestään.
DMEM, joka on johdettu Eagle's Minimal Essential Mediumista (EMEM), tarjoaa edeltäjäänsä suuremman pitoisuuden aminohappoja ja vitamiineja. Sen sijaan Ham's F-12 perustuu Ham's F-10 -alustaan ja tarjoaa täydentävän joukon välttämättömiä komponentteja.
Optimaalisen solujen kasvun tukemiseksi on yleistä täydentää DMEM:Ham's F12:ta FBS:llä tyypillisesti 5–10 %:n pitoisuudella. Tämä lisäys on tarpeen, koska väliaineesta puuttuu solujen kehitykselle välttämättömiä kasvuhormoneja, lipidejä ja proteiineja.
DMEM:Ham's F12 sisältää pH-puskuriin ja siihen lisätään usein fenolipunainen, joka on pH-indikaattori. DMEM:Ham's F12:ssa tai missä tahansa bikarbonaattipuskuriin perustuvassa elatusaineessa viljellyt solut vaativat 5–10 %:n CO2-pitoisuuden, jotta sopiva pH-taso säilyy.
Laadunvalvonta
Steriilisuodatettu
Säilytys ja säilyvyysaika
Säilytä +2–+8 °C:ssa, suojattuna valolta.
Avatun pakkauksen säilytyslämpötila on 4 °C, ja se on käytettävä 6–8 viikon kuluessa.
Kuljetusolosuhteet
Ympäristön lämpötila
Säilytys
Säilytä jääkaapissa +2 °C – +8 °C:ssa pimeässä. Vältä jäädyttämistä ja toistuvaa lämmittämistä +37 °C:seen, sillä se heikentää tuotteen laatua.
Älä lämmitä elatusainetta yli 37 °C:n lämpötilaan tai käytä hallitsemattomia lämmönlähteitä, kuten mikroaaltouuneja.
Jos vain osa elatusaineesta käytetään, ota tarvittava määrä ja lämmitä se huoneenlämpötilaan ennen käyttöä.
Koostumus
Luokka
Komponentit
Pitoisuus (mg/l)
Aminohapot
Glysiini
18,75
L-alaniini
4,45
L-arginiini-HCl
147,50
L-asparagiini H₂O
7,50
L-asparagiinihappo
6,65
L-kysteiini-HCl-H₂O
17,56
L-kystiini 2 HCl
31,29
L-glutamiinihappo
7,35
L-glutamiini
365,00
L-histidiini-HCl-H₂O
31,48
L-isoleusiini
54,47
L-leusiini
59,05
L-lysiini-HCl
91,25
L-metioniini
17,24
L-fenyylialaniini
35,48
L-proliini
17,25
L-seriini
26,25
L-treoniini
53,45
L-tryptofaani
9,02
L-tyrosiini 2 Na 2 H2O
55,79
L-valiini
52,85
Vitamiinit
D-biotiini
0,0035
Koliinikloridi
8,98
D-kalsiumpantotenaatti
2,24
Foolihappo
2,66
myo-inositoli
12,60
Nikotiiniamidi
2,02
Pyridoksiini-HCl
0,031
Pyridoksaali-HCl
2,00
Riboflaviini
0,219
Tiamiini-HCl
2,17
B12-vitamiini
0,68
Epäorgaaniset suolat
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO3)3 9 H2O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Muut komponentit
D-glukoosi
3151,00
Hypoxantiini
2,40
HEPES
3574,50
Linolihappo
0,042
Lipoiinihappo
0,105
Fenolipunaisen natriumsuola
8,63
Putreskiini-2-HCl
0,081
Natriumpyruvaatti
55,00
Tymidiini
0,365
, w: 3,0 g/l glukoosia, w: vakaa glutamiini, w: 2,0 mM natriumpyruvaattia, w: 2,2 g/l NaHCO3")
- 0,02 lämpötilassa 20–25 °C. Jokainen erä on testattu steriiliyden ja mykoplasman ja bakteerien puuttumisen osalta. Säilytys Säilytä jääkaapissa +2–+8 °C:ssa pimeässä. Jäädyttäminen ja lämmittäminen +37 °C:seen heikentää tuotteen laatua. Älä lämmitä väliainetta yli 37 °C:seen tai käytä hallitsemattomia lämmönlähteitä (esim. mikroaaltouunit). Jos vain osa väliaineesta on tarkoitus käyttää, ota tarvittava määrä pullosta ja lämmitä se huoneenlämpötilaan. Kaikkien väliaineiden, paitsi perusväliaineen, säilyvyysaika on 6–8 viikkoa avaamispäivästä. Koostumus Komponentit mg/l Epäorgaaniset suolatKalsiumkloridi x 2H2O132,0 Magnesiumsulfaatti97,67 Kaliumkloridi400 Natriumkloridi6 460,00 Dinatriumvetyfosfaatti (vedetön)504,00 Muut komponentitD(+)-glukoosi (vedetön)3 000 Glutationi (pelkistetty)0 Lihapeptoni600 Fenolipunainen natriumsuola11 AminohapotL-alaniini13 L-arginiini x HCl42,1 L-asparagiini x H2O45,03 L-asparagiinihappo19,97 L-kysteiini x HCl x H2O31,75 L-glutamiini (stabiili)219,15 L-glutamiinihappo22,07 Glysiini7,5 L-histidiini x HCl x H2O20,96 L-hydroksiproliini19,67 L-isoleusiini39,3 L-leusiini39,36 L-lysiini x HCl36,54 L-metioniini14,92 L-fenyylialaniini16,52 L-proliini17,27 L-seriini26,28 L-treoniini17,87 L-tryptofaani3,06 L-tyrosiinin dinatriumsuola26,1 L-valiini17,57 Vitamiinitp-Aminobentsoehappo1,0 Askorbiinihappo0,5 D(+)-biotiini0,2 D-kalsiumpantotenaatti0 Koliinikloridi5 Foolihappo10 Myo-inositoli36 Nikotinamidi0 Nikotiinihappo0,5 Pyridoksaali-HCl0,5 Pyridoksiini-HCl0,5 Riboflaviini0,2 Tiamiini-HCl0 B12-vitamiini2,0

Medium 199 tarjoaa monenlaisia sovelluksia alalla. Sillä voidaan tehokkaasti ylläpitää kumulaari
- ja munasolukompleksia (COC) ja tukea munasolujen in vitro -kypsymistä. Lisäksi sitä käytetään saksalaisilta holsteinlehmiltä kerättävien munasolujen huuhtelussa. Medium 199 on lisäksi erinomainen väliaine rotista peräisin olevien sydämen endoteelisolujen viljelyyn. Nämä sovellukset osoittavat Medium 199:n monipuolisuuden ja mukautuvuuden erilaisiin kokeellisiin tarpeisiin.
Historia
Medium 199:n kehittäminen 1950-luvulla merkitsi merkittävää edistystä kudosviljelyaineissa. Ennen sen käyttöönottoa monet kasvatusmediat perustuivat eläimistä peräisin oleviin tuotteisiin ja kudosuutteisiin. Morgan ja kollegat kuitenkin mullistivat alan muotoilemalla täysin määritellyn ravinnelähteen soluviljelmille. Kokeilemalla erilaisia vitamiinien, aminohappojen ja muiden tekijöiden yhdistelmiä he havaitsivat Medium 199:n poikkeukselliset kasvua edistävät ominaisuudet.
Laadunvalvonta
pH = 7,2 +/
- 0,02 20-25 °C:ssa.
Jokainen erä on testattu steriiliyden sekä mykoplasman ja bakteerien puuttumisen varalta.
Ylläpito
Säilytetään jääkaapissa +2°C
- +8°C:ssa pimeässä. Pakastaminen ja lämmittäminen +37° C:een asti minimoi tuotteen laadun.
Älä lämmitä elatusainetta yli 37° C:seen tai käytä hallitsemattomia lämmönlähteitä (esim. mikroaaltolaitteita).
Jos vain osa väliaineesta on tarkoitus käyttää, poista tämä määrä pullosta ja lämmitä se huoneenlämmössä.
Kaikkien elatusaineiden, lukuun ottamatta peruselatusainetta, säilyvyysaika on 8 viikkoa valmistuspäivästä.
Koostumus
Ainesosat
mg/l
Epäorgaaniset suolat
Kalsiumkloridi x 2H2O
264,92
Rauta(III)nitraatti x 9H2O
0,72
Magnesiumsulfaatti
97,67
Kaliumkloridi
400,00
Natriumasetaatti x 3H2O
82,95
Natriumkloridi
6,800.00
Natriumdivetyfosfaatti x H2O
140,00
Muut komponentit
Adeniinisulfaatti
10,00
AMP
0,20
ATP
1,00
Kolesteroli
0,20
2'-Deoksiriboosi
0,50
D(+)-glukoosi vedetön
1,000.00
Glutationi (punainen)
0,05
Guaniini x HCl
0,30
Hypoksantiini
0,30
Fenolipunainen
10,00
D-Riboosi
0,50
Timiini
0,30
Tween 80
4,90
Uracil
0,30
Ksantiini
0,30
NaHCO3
2,200.00
Aminohapot
L-alaniini
25,00
L-arginiini x HCl
70,00
L-asparagiinihappo
30,00
L-kysteiini x HCl x H2O
0,10
L-kystiini
20,00
L-Glutamiini vakaa
149,00
L-glutamiinihappo
67,00
Glysiini
50,00
L-histidiini x HCl x H2O
21,88
L-hydroksiproliini
10,00
L-isoleusiini
20,00
L-leusiini
60,00
L-lysiini x HCl
70,00
L-metioniini
15,00
L-Fenyylialaniini
25,00
L-proliini
40,00
L-Seriini
25,00
L-treoniini
30,00
L-tryptofaani
10,00
L-tyrosiini
40,00
L-valiini
25,00
Vitamiinit
4-Aminobentsoehappo
0,05
Askorbiinihappo
0,05
D(+)-Biotiini
0,01
Kalsiferoli
0,10
D-kalsiumpantotenaatti
0,01
Koliinikloridi
0,50
Foolihappo
0,01
myo-Inositoli
0,05
Menadioni
0,01
Nikotiinihappo
0.025
Nikotiiniamidi
0.025
Pyridoksaali x HCl
0.025
Pyridoksoli x HCl
0.025
Riboflaviini
0,01
DL-α-tokoferolifosfaatin dinatriumsuola
0,01
Tiamiini x HCl
0,01
A-vitamiini asetaatti
0,14

IMDM soveltuu hyvin nopeasti lisääntyville, suuren tiheyden soluviljelmille, kuten Jurkat-, COS-7
- ja makrofagisoluille. IMDM:n eri modifikaatiot, joita on saatavilla erilaisiin soluviljelysovelluksiin, löytyvät mediavalintatyökalun avulla. Nestemäiset väliaineet tarjoavat välttämättömiä ravintoaineita kaikkiin soluviljelysovelluksiin. Jokainen korkealaatuinen soluviljelymediamme valmistetaan alun perin julkaistun kaavan tai sen modifikaatioiden mukaisesti, jotka ovat tarpeen viljelymedian tasaisen suorituskyvyn ja vakauden varmistamiseksi.
IMDM vs. DMEM
IMDM sisältää kaliumnitraattia rautanitraatin sijasta sekä HEPES:ää ja natriumpyruvaattia. IMDM:n sisältämien lisäkomponenttien ansiosta se soveltuu paremmin erikoistuneisiin solutyyppeihin ja erityissovelluksiin kuin DMEM.
IMDM vs. RPMI
IMDM:llä ja RPMI:llä on erilaiset koostumukset, joilla voi olla merkitystä PMA/ionomysiini-stimulaation kannalta. Yksi merkittävä ero on Ca2+ -pitoisuus. RPMI sisältää 0,42 mM Ca2+:a, kun taas IMDM sisältää 1,49 mM Ca2+:a.
Laadunvalvonta
pH = 7,2 +/
- 0,02 20-25 °C:ssa.
Jokainen erä on testattu steriiliyden sekä mykoplasman ja bakteerien puuttumisen varalta.
Ylläpito
Säilytetään jääkaapissa +2°C
- +8°C:ssa pimeässä. Pakastaminen ja lämmittäminen +37° C:een asti minimoi tuotteen laadun.
Älä lämmitä elatusainetta yli 37° C:seen tai käytä hallitsemattomia lämmönlähteitä (esim. mikroaaltolaitteita).
Jos vain osa väliaineesta on tarkoitus käyttää, poista tämä määrä pullosta ja lämmitä se huoneenlämmössä.
Kaikkien elatusaineiden, lukuun ottamatta peruselatusainetta, säilyvyysaika on 8 viikkoa valmistuspäivästä.
Koostumus
Ainesosat
mg/l
Epäorgaaniset suolat
Kalsiumkloridi x 2 H2O
219,00
Kaliumkloridi
330,00
Kaliumnitraatti
0.076
Vedetön magnesiumsulfaatti
97,73
Natriumkloridi
4,505.00
Natriumdivetyfosfaatti vedetön
109,00
Natriumseleniitti
0,02
Muut komponentit
D(+)-glukoosi vedetön
4,500.00
HEPES
5,958.00
Natriumpyruvaatti
110,00
Fenolipunainen
15,00
Aminohapot
L-alaniini
25,00
L-arginiini x HCl
84,00
L-asparagiini x H2O
25,00
L-asparagiinihappo
30,00
L-kystiini x 2HCl
91,24
L-Glutamiini
584,00
L-glutamiinihappo
75,00
Glysiini
30,00
L-histidiini x HCl x H2O
42,00
L-isoleusiini
104,80
L-leusiini
104,80
L-lysiini x HCl
146,20
L-metioniini
30,00
L-Fenyylialaniini
66,00
L-proliini
40,00
L-Seriini
42,00
L-treoniini
95,20
L-tryptofaani
16,00
L-tyrosiini x 2Na
104,20
L-valiini
93,60
Vitamiinit
D(+)-Biotiini
0.013
D-kalsiumpantotenaatti
4,00
Koliinikloridi
4,00
Foolihappo
4,00
myo-Inositoli
7,20
Soluviljelyalustat: yleiskatsaus
Elämäntieteiden alalla yksi tärkeimmistä menetelmistä on soluviljely. Soluviljelyllä tarkoitetaan solujen, kudosten tai elinten poistamista eläimestä tai kasvista ja niiden siirtämistä keinotekoiseen ympäristöön, joka on suotuisa niiden selviytymiselle ja/tai kasvulle. Solujen optimaalisen kehityksen kannalta olennaisia ympäristötekijöitä ovat säädelty lämpötila, solujen kiinnittymiselle sopiva substraatti, asianmukainen kasvualusta sekä inkubaattori, joka ylläpitää optimaalista pH-arvoa ja osmolaliteettia. Solujen on saatava nämä olosuhteet, jotta ne voivat kasvaa täyteen potentiaaliinsa.
Sopivan kasvualustan valinta in vitro -viljelyä varten on soluviljelyn kriittisin ja samalla tärkein vaihe. Kasvualusta, jota kutsutaan myös viljelyalustaksi, on neste tai geeli, joka on muotoiltu edistämään organismien kehitystä mikroskooppisella, solutasolla tai kasvien kaltaisella tasolla. Solujen viljelyyn käytetty alusta sisältää usein riittävän määrän energiaa ja aineita, jotka säätelevät solusykliä. Viljelyalustan pääkomponentteja ovat aminohapot, vitamiinit, epäorgaaniset suolat, glukoosi ja seerumi. Seerumi lisätään alustaan, koska se toimii kasvutekijöiden, hormonien ja kiinnittymistekijöiden lähteenä. Ravinteiden tarjoamisen lisäksi alusta auttaa myös ylläpitämään pH- ja osmolaliteettitasoja.
Soluviljelyssä käytettävät elatusainetyypit
Sekä ihmisen että eläinten soluja voidaan kasvattaa joko keinotekoisessa tai synteettisessä elatusaineessa tai täysin luonnollisessa elatusaineessa, jota on täydennetty luonnollisilla aineilla. Seuraavassa esittelemme yleiskatsauksen nykyisin saatavilla olevista elatusainetyypeistä.
Luonnolliset elatusaineet
Luonnollisissa elatusaineissa on ainoastaan luonnollisessa tilassaan esiintyviä biologisia nesteitä. Luonnolliset elatusaineet ovat erittäin hyödyllisiä ja helppokäyttöisiä monenlaisten eläinsolutyyppien viljelyssä. Luonnollisten elatusaineiden tarkkojen koostumusten puutteellinen ymmärtäminen on tärkein tekijä, joka vaikuttaa luonnollisilla elatusaineilla saatujen tulosten heikkoon toistettavuuteen.
Keinotekoiset elatusaineet
Keinotekoisten tai synteettisten elatusaineiden valmistukseen kuuluu ravinteiden (sekä orgaanisten että epäorgaanisten), seerumiproteiinien, hiilihydraattien, kofaktoreiden, vitamiinien ja suolojen sekä O2- ja CO2-kaasuvaiheiden lisääminen [1].
Erilaisia keinotekoisia elatusaineita on kehitetty täyttämään yksi tai useampi seuraavista tehtävistä: 1) Välitön eloonjääminen (tasapainoinen suolaliuos, jonka pH ja osmoottinen paine on tarkasti säädetty). 2) Pitkäaikainen eloonjääminen (tasapainoinen suolaliuos, johon on lisätty erilaisia orgaanisten kemikaalien ja/tai seerumin yhdistelmiä). 3) Määrittelemätön kehitys. 4) Erikoistoiminnot.
Keinotekoiset elatusaineet luokitellaan neljään ryhmään:
Seerumia sisältävät elatusaineet
Eläinsolujen viljelyyn käytettävissä elatusaineissa yleisimmin käytetty lisäaine on naudan sikiöseerumi. Sitä lisätään viljelyalustaan edullisena lisäaineena parhaiden mahdollisten kasvuolosuhteiden saavuttamiseksi. Sen lisäksi, että se toimii epästabiilien tai veteen liukenemattomien ravintoaineiden, hormonien ja kasvutekijöiden, proteaasi-inhibiittorien ja muiden aineiden kuljettajana tai kelatoijana, seerumi myös sitoo ja neutraloi haitallisia molekyylejä.
Seerumittomat elatusaineet
Seerumin läsnäololla elatusaineessa on useita haittoja, ja se voi aiheuttaa merkittäviä tulkintavirheitä immunologisessa tutkimuksessa [2, 3]. On kehitetty useita erilaisia seerumittomia elatusaineita [4, 5]. Nämä elatusaineet on yleensä kehitetty nimenomaan tukemaan yhden solutyypin viljelyä, kuten Thermo Fisher Scientificin Knockout Serum Replacement ja Knockout DMEM sekä Stem Cell Technologiesin mTESR-elatusaine [6], joka on tarkoitettu kantasoluille [7].
Lisäksi näihin elatusaineisiin on lisätty määriteltyjä määriä puhdistettuja kasvutekijöitä, lipoproteiineja ja muita proteiineja, joita muuten tyypillisesti saadaan seerumista [8]. Näitä elatusaineita kutsutaan usein ”määritellyiksi elatusaineiksi”, koska niiden koostumuksen komponentit tunnetaan hyvin.
Kemiallisesti määritellyt elatusaineet
Näihin elatusaineisiin kuuluu erittäin puhtaita epäorgaanisia ja orgaanisia komponentteja, joita ei ole saastuttanut minkäänlainen kontaminaatio. Niihin voi kuulua myös puhtaita proteiineja, kuten kasvutekijöitä.
Bakteerien tai hiivan geneettinen muuntaminen sekä tiettyjen rasvahappojen, vitamiinien, kolesterolin ja aminohappojen lisääminen johtaa niiden komponenttien tuotantoon [9].
Proteiinittomat elatusaineet
Proteiinittomat elatusaineet ovat sellaisia, jotka eivät sisällä lainkaan proteiinia, vaan ainoastaan muita kuin proteiini-aineosia. Verrattuna elatusaineisiin, joihin on lisätty seerumia, proteiinittomien elatusaineiden käyttö edistää solujen voimakkaampaa lisääntymistä ja proteiinin ilmentymistä sekä helpottaa loppuprosessissa syntyvän tuotteen puhdistamista [10–12]. Proteiinia ei sisälly esimerkiksi MEM- ja RPMI-1640-valmisteisiin. Proteiinilisää voidaan kuitenkin antaa, jos se on tarpeen.
Viljelyalustat ja niiden peruskomponentit
Kaupallisia viljelyalustoja on saatavana jauheena tai nesteenä, ja ne sisältävät usein erilaisia ravintoaineita, kuten aminohappoja, glukoosia, suoloja, vitamiineja ja muita ravintolisäaineita.
Näiden komponenttien tarve vaihtelee solulinjoittain, ja nämä vaihtelut selittävät viljelyalustojen laajan valikoiman. Jokaisella komponentilla on tietty tehtävä, jota kuvataan seuraavissa kappaleissa:
Puskurointijärjestelmät
Optimaalisten kasvuolosuhteiden ylläpitämiseksi pH:ta on säädettävä, mikä tapahtuu usein jommankumman seuraavista puskurointijärjestelmistä:
Luonnollinen puskurointijärjestelmä
Ilmakehän CO2/H2CO3-suhde on sama kuin elatusaineessa, mikä luo luonnollisen puskurointimekanismin. Luonnollisen puskurointimekanismin säilyttämiseksi viljelmät on pidettävä ilmakehässä, jossa CO2-pitoisuus on 5–10 %, mikä saavutetaan usein käyttämällä CO2-inkubaattoria. Yksi luonnollisen puskurin suurimmista eduista on sen edullisuus ja turvallisuus.
HEPES
Zwitterionia HEPES:ää käyttävä kemiallinen puskurointi on tehokkaampaa pH-alueella 7,2–7,4 eikä vaadi säädeltyä kaasuympäristöä. Tietyille solutyypeille suurempi HEPES-annos voi olla haitallista. HEPES:ää sisältävät elatusaineet ovat myös huomattavasti alttiimpia fluoresoivan valon fototoksisille vaikutuksille [13].
Fenolipunainen
pH-indikaattori fenolipunainen sisältyy usein kaupallisesti saatavilla oleviin viljelyalustoihin, mikä mahdollistaa pH:n jatkuvan seurannan. Solujen lisääntyessä niiden tuottamat metaboliitit aiheuttavat pH:n muutoksen ja siten alustan värinmuutoksen. Fenolipunaisella on kaksijakoinen vaikutus elatusaineen väriin: se muuttuu keltaiseksi happamassa pH:ssa ja purppuraksi emäksisessä pH:ssa. Soluviljelylle optimaalinen arvo, pH 7,4, saa elatusaineen näyttämään fluoresoivasti punaiselta.
Fenolipunaisella on kuitenkin muutamia haittoja: Ensinnäkin fenolipunainen kykenee jäljittelemään useiden steroidihormonien, pääasiassa estrogeenin, toimintaa [14]. Siksi tutkittaessa estrogeenille herkkiä soluja, kuten rintakudosta, suositellaan fenolipunaisesta vapaata elatusainetta. Fenolipunainen häiritsee natrium-kalium-tasapainoa useissa seerumittomissa valmisteissa. Tämän vaikutuksen voi kumota lisäämällä elatusaineeseen seerumia tai naudan aivolisäkkeen hormonia [15]. Kolmanneksi fenolipunainen haittaa havaitsemista virtaussytometrisissä kokeissa.
Epäorgaaniset suolat
Epäorgaanisia suoloja, kuten natrium-, kalium- ja kalsiumioneja, sisältävät elatusaineet auttavat ylläpitämään osmoottista tasapainoa ja säätelemään kalvopotentiaalia.
Aminohapot
Koska aminohapot ovat proteiinin peruskomponentteja, ne ovat olennainen osa jokaista koskaan kehitettyä solujen kasvualustaa. Koska solut eivät kykene tuottamaan tiettyjä aminohappoja itse, on tärkeää, että viljelyalustassa on välttämättömiä aminohappoja. Ne ovat välttämättömiä solujen lisääntymiselle, ja niiden pitoisuus määrää saavutettavissa olevan suurimman solutiheyden. Erityisesti välttämätön aminohappo L-glutamiini on erittäin tärkeä.
L-glutamiini toimii aineenvaihdunnan toissijaisena energialähteenä ja toimittaa typpeä NAD:n, NADPH:n ja nukleotidien tuotantoon. Koska L-glutamiini on epästabiili aminohappo, joka ajan myötä muuttuu muotoon, jota solut eivät pysty hyödyntämään, se on lisättävä elatusaineeseen.
Lisäksi kasvualustaan voidaan lisätä ei-välttämättömiä aminohappoja korvaamaan kasvun aikana kuluneita aminohappoja. Solujen kasvu kiihtyy ja niiden elinkelpoisuus paranee, kun kasvualustaa täydennetään ei-välttämättömillä aminohapoilla.
Hiilihydraatit
Sokereina esiintyvät hiilihydraatit ovat tärkein energialähde. Monissa elatusaineissa on glukoosin ja galaktoosin lisäksi myös maltoosia ja fruktoosia.
Proteiinit ja peptidit
Albumiini, transferriini ja fibronektiini ovat yleisimmin käytettyjä proteiineja ja peptidejä. Ne ovat erityisen tärkeitä elatusaineissa, jotka eivät sisällä seerumia. Albumiini, transferriini, aprotiniini, fetuiini ja fibronektiini ovat joitakin proteiineja, joita voi esiintyä seerumissa, joka on runsas proteiinin lähde.
Albumiini on veren pääasiallinen proteiini, ja sen tehtävänä on sitoa ja kuljettaa erilaisia aineita, kuten vettä, suoloja, vapaita rasvahappoja, hormoneja ja vitamiineja, eri elinten ja solujen välillä. Albumiinin kyky sitoutua kemikaaleihin tekee siitä tehokkaan vaihtoehdon haitallisten yhdisteiden poistamiseksi soluviljelyalustasta.
Aprotiniini on suoja-aine soluviljelyjärjestelmissä, sillä se on stabiili neutraalissa ja happamassa pH:ssa sekä kestää korkeita lämpötiloja ja proteolyyttisten entsyymien mahdollisesti aiheuttamaa hajoamista. Se kykenee estämään useita seriini-proteaaseja, muun muassa trypsiiniä.
Fetuini on glykoproteiini, jota voi esiintyä suurempina pitoisuuksina sikiöiden ja vastasyntyneiden eläinten seerumissa verrattuna aikuisten seerumiin. Lisäksi se toimii seriiniproteaasi-inhibiittorina. Fibronektiini-proteiini on olennainen osa solujen adheesioprosessia. Transferriini on rautaa kuljettava proteiini, joka vastaa raudan kuljettamisesta solujen kalvoihin.
Rasvahapot ja lipidit
Niillä on ratkaiseva rooli seerumittomassa elatusaineessa, kun seerumia ei ole läsnä.
Vitamiinit
Lukuisat vitamiinit ovat välttämättömiä solujen kehitykselle ja lisääntymiselle. Solut eivät pysty tuottamaan vitamiineja riittävissä määrissä, joten ne ovat välttämättömiä kudosviljelyssä ravintolisinä.
Soluviljelyssä seerumi on vitamiinien pääasiallinen lähde; viljelyalustoja kuitenkin rikastetaan myös erilaisilla vitamiineilla, jotta ne sopisivat tietylle solutyypille. Kasvun stimuloimiseen käytetään tyypillisimmin B-ryhmän vitamiineja.
Hivenaineet
Kemiallisia alkuaineita, kuten kuparia, sinkkiä, seleeniä ja trikarboksyylihapon välituotteita, kutsutaan hivenaineiksi. Hivenaineita lisätään usein seerumittomiin elatusaineisiin korvaamaan niitä, joita tyypillisesti esiintyy seerumissa. Nämä alkuaineet ovat tärkeitä kemiallisia komponentteja, joita tarvitaan terveen solun kehitykseen. Monet biokemialliset reaktiot, kuten entsyymien toiminta, riippuvat tietyistä mikroravinteista.
Elatusaineiden lisäaineet
Tiettyjen solulinjojen kasvulle suositeltava täydellinen kasvualusta vaatii lisäkomponentteja, joita ei ole peruskasvualustassa eikä seerumissa. Nämä ravintolisät tukevat solujen kasvua ja asianmukaista aineenvaihduntaa.
Vaikka hormonit, kasvutekijät ja signalointimolekyylit ovat välttämättömiä tiettyjen solulinjojen asianmukaiselle lisääntymiselle, seuraavia varotoimia on aina noudatettava: Koska lisäaineiden lisääminen saattaa muuttaa täydellisen kasvualustan osmolaliteettia, mikä voi estää solujen kehitystä, on aina suositeltavaa tarkistaa osmolaliteetti lisäaineiden lisäämisen jälkeen. Useimmille solulinjoille optimaalinen osmolaliteetti on välillä 260–320 mOSM/kg.
Antibiootit
Antibiootteja käytetään usein bakteeri- ja sienikontaminaatioiden estämiseen [16], vaikka ne eivät ole välttämättömiä solujen kasvulle. Koska antibiootit voivat peittää mykoplasma- ja resistenttien bakteerien aiheuttaman kontaminaation, niiden rutiinikäyttöä ei suositella soluviljelyssä [17, 18].
Lisäksi antibiootit voivat häiritä yliherkkien solujen aineenvaihduntaa. Usein käytetään MilliporeSigman ja Life Technologiesin valmistamia penisilliini-streptomysiini-yhdistelmiä. Plasmocinia on käytetty gliomasolulinjojen TS603, TS516 ja BT260 viljelyssä [19], ja sen on osoitettu olevan tehokas mykoplasmakontaminaation poistamisessa (20).
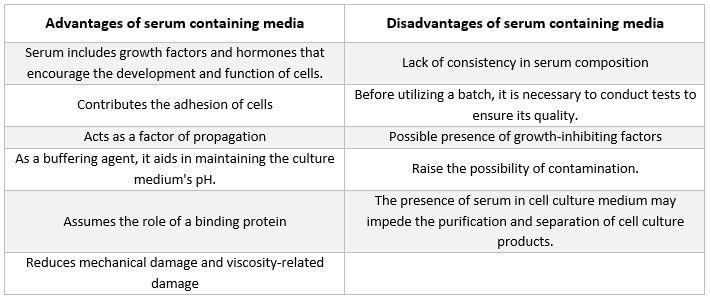
Seerumi
Seerumissa on albumiineja, kasvutekijöitä ja kasvun estäjiä. Seerumi on yksi soluviljelyalustan merkittävimmistä komponenteista, koska se tarjoaa aminohappoja, proteiineja, vitamiineja (erityisesti rasvaliukoisia vitamiineja, kuten A, D, E ja K), hiilihydraatteja, lipidejä, hormoneja, kasvutekijöitä, mineraaleja ja hivenaineita.
Sikiön ja vasikan seerumia käytetään usein viljeltyjen solujen kehityksen edistämiseen. Sikiön seerumi sisältää runsaasti kasvutekijöitä ja sopii solujen kloonaukseen sekä herkkien solujen kehittämiseen. Koska vasikan seerumin kasvua edistävät ominaisuudet ovat heikentyneet, sitä käytetään kontaktinestokokeissa. Tavalliset kasvualustat sisältävät usein 2–10 % seerumia. Seerumin lisääminen viljelyalustaan palvelee seuraavia tarkoituksia [21]:
-
Seerumi toimittaa soluille välttämättömiä ravintoaineita (sekä liuoksessa että proteiineihin sitoutuneina).
-
Seerumissa on useita kasvutekijöitä ja hormoneja, jotka osallistuvat kasvun edistämiseen ja solujen erityistoimintaan.
-
Se tarjoaa monia sitoutuvia proteiineja, kuten albumiinia ja transferriiniä, jotka kuljettavat muita kemikaaleja soluun. Esimerkiksi albumiini kuljettaa rasvoja, vitamiineja, hormoneja jne. soluihin.
-
Se tarjoaa myös proteiineja, kuten fibronektiiniä, jotka lisäävät solujen tarttumista substraattiin. Lisäksi se tuottaa leviämiselementtejä, jotka auttavat solujen laajenemisessa ennen jakautumista.
-
Se toimittaa proteaasi-inhibiittoreita, jotka estävät proteolyysin soluissa.
-
Se sisältää myös mineraaleja, kuten Na+, K+, Zn2+ ja Fe2+.
-
Se lisää väliaineen viskositeettia ja suojaa siten soluja mekaanisilta vaurioilta suspensiokulttuurin sekoituksen aikana.
-
Se toimii myös puskurina.
Viitteet
[1] Morgan J, Morton H, Parker R. Eläinsolujen ravitsemus kudosviljelyssä; alustavia tutkimuksia synteettisestä elatusaineesta. Proc Soc Exp Biol Med. 1950;73:1–8
[2] Kerbel R, Blakeslee D. Sikiön vasikan seerumin komponentin nopea adsorptio viljellyissä nisäkässoluissa. Mahdollinen artefaktien lähde solukohtaisia antigeenejä vastaan suunnattujen antiseerumien tutkimuksissa. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. Seerumin lisääminen solususpensioiden valmistuksessa käytettyyn elatusaineeseen mahdollisena artefaktien lähteenä polvitaipeen imusolmuketestillä tutkituissa soluvälitteisissä reaktioissa. J Immunogenet. 1980;7:483–9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Kaupalliset seerumittomat elatusaineet: hybridomien kasvatus ja monoklonaalisten vasta-aineiden tuotanto. J Immunol Methods. 1991;145:175–83
[5] Barnes D, Sato G. Menetelmät viljeltyjen solujen kasvattamiseksi seerumittomassa elatusaineessa. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O, ym. HSP70-kaperonit kuljettavat RNA:ta sisältämättömän TDP-43:n anisotrooppisiin, ytimen sisäisiin nestemäisiin pallomaisiin kuoriin. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, ym. Downin oireyhtymän aiheuttama ikääntyminen häiritsee hermosolujen esisolujen tumarakennetta. Cell Stem Cell. 2022;29:116–130.e7
[8] Iscove N, Melchers F. Seerumin täydellinen korvaaminen albumiinilla, transferriinillä ja soijalipidillä lipopolysakkaridille reagoivien B-lymfosyyttien viljelmissä. J Exp Med. 1978;147:923–33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Hybridomien kasvua ja monoklonaalisten vasta-aineiden tuotantoa varten kehitetyn kemiallisesti määritellyn proteiinittoman elatusaineen järjestelmällinen parantaminen. J Biotechnol. 1996;45:111–23
[10] Darfler F. Proteiiniton elatusaine hybridomien ja muiden immuunijärjestelmän solujen kasvattamiseen. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Seerumiton soluviljely: yhtenäistävä lähestymistapa. Cell. 1980;22:649–55
[12] Hamilton W, Ham R. Kiinalaisten hamsterisolulinjojen klonaalinen kasvatus proteiinittomissa elatusaineissa. In Vitro. 1977;13:537–47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Valolle altistetun HEPES-pitoisen viljelyväliaineen sytotoksisten vaikutusten analyysi. In Vitro Cell Dev Biol. 1985;21:282–7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. Fenolipunainen kudosviljelyalustoissa on heikko estrogeeni: vaikutukset estrogeenille reagoivien solujen viljelytutkimukseen. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Seerumittomien elatusaineiden kehittäminen. Teoksessa: Master JRW, toim. Animal Cell culture, 3. painos. Oxford: Oxford University Press; 2000.
[16] Perlman D. Antibioottien käyttö soluviljelyalustoissa. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. Soluviljelmien mykoplasma-infektioiden leviäminen ja torjunta. In Vitro. 1976;12:643–8
[18] Masters J, Stacey G. Elatusaineen vaihtaminen ja solulinjojen siirrostaminen. Nat Protoc. 2007;2:2276–84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M, ym. Histonidemetylaasi KDM6A aistii suoraan happea säätääkseen kromatiinia ja solujen kohtaloa. Science. 2019;363:1217–1222
[20] Molla Kazemiha V, Azari S, Amanzadeh A, Bonakdar S, Shojaei Moghadam M, Habibi Anbouhi M, ym. Plasmocin™:n tehokkuus erilaisissa mollicute-bakteerien tartuttamissa nisäkässolulinjoissa verrattuna soluviljelyssä yleisesti käytettyihin antibiootteihin: paikallinen kokemus. Cytotechnology. 2011;63:609-20
[21] Kragh Hansen U. Ligandin sitoutumisen seerumialbumiiniin molekyylitason näkökohdat. Pharmacol Rev. 1981;33:17–53