
HepG2-rakud – maksavähi uurimise ressurss
Hep-G2 on inimese maksavähi rakuliin, mis pärineb 15-aastase kaukaasia päritolu mehe maksakoe rakkudest, kellel oli hepatotsellulaarne kartsinoom. Neid rakke kasutatakse sageli ravimite metabolismi ja hepatotoksilisuse uuringutes. Kuigi HepG2 rakkudel on kõrge proliferatsioonikiirus ja epiteelilaadne välimus, ei ole need tuumorigeensed ning täidavad mitmesuguseid diferentseeritud maksa funktsioone. 1975. aastal eraldasid teadlased HepG2 rakud hepatotsellulaarsest kartsinoomist, muutes need esimeseks maksarakuliiniks, millel esinevad hepatotsüütide olulised omadused. Erinevalt varem loodud SK-Hep1 rakuliinist, millel puuduvad olulised maksarakkude markerid, suudavad HepG2 rakud eritada mitmesuguseid plasmaproteiine ja pakuvad väärtuslikku mudelit inimese hepatotsüütide rakupinna domeenide rakusisese dünaamika uurimiseks. Need rakud näitavad epiteelilaadset morfoloogiat, nende kromosoomide arv on tavaliselt 55 ja neid saab stimuleerida inimese kasvuhormooniga.
- Kasvukeskkond
- Vaata tootelehte
- Kahekordistumisaeg
- Vaata tootelehte
- Kasvutüüp
- Adherent
- Bioloogiline ohutustase
- BSL-1
- Saadaval
- Cytion — Telli HepG2
HepG2 omadused
Esmaste hepatotsüütide tüüpiline kuju on kuubikujuline ja neis on tavaliselt kaks tuuma. Seevastu HepG2 rakkudel on epiteelilaadne morfoloogia, neis on üks tuum ja kromosoomide arv on 48–54 rakku kohta. Kuigi HepG2 rakud võivad moodustada kuni 25% kogu raku valgust, on nende suurus suurem kui normaalse hepatotsüüdi oma, moodustades umbes 10% raku kogu valgust. Rakuvalkud on rakus olulised tegurid, täites geenide poolt määratud funktsioone.
Tuumorrakud, sealhulgas need, millel on ebanormaalne kromosoomide arv, näitavad sageli tuumade arvu suurenemist, kuni seitsme tuumani rakus. Tänu oma kõrge diferentseerumisastmele in vitro on HepG2-rakud ideaalne mudel inimese hepatotsüütide sisesele transportimisele ja sapikanalite, sinusoidide membraanivalkude ning lipiidide dünaamikale uurimiseks.
HepG2-raku keskmine läbimõõt on umbes 10–20 µm, mis on väiksem kui hepatotsüüdi läbimõõt (15 µm), kuid sarnane hepatoblastoomi (HB) kasvajarakkudega, mille läbimõõt on 10–20 µm.
HepG2 geneetika
Hep-G2 rakuliinil esineb mitmeid translokatsioone, sealhulgas kromosoomide 1 ja 21 lühikeste harude vahelisi, kromosoomide 2, 16 ja 17 trisoomiaid ning kromosoomi 20 tetrasoomiat. Samuti on täheldatud kromosoomi 4q3 piirkonna kadu, mis on seotud translokatsiooniga t(1;4), mida esineb sageli hepatoblastoomis (HB), ning muude kromosoomianomaaliatega, nagu trisoomiad 2 ja 20. HepG2-rakkude kromosoomide arv ulatub 50-st 60-ni, mis viitab hüperdiploidsele kariotüübile, samas kui mõnel juhul esineb üle 100 kromosoomi ja neid iseloomustab tetraploidsus. HepG2 rakud sisaldavad umbes 7,5 pg DNA-d, mis on 15% rohkem kui keskmisel somaatilisel rakul. Võrdluseks: esmased hepatotsüüdid on kuubikujulised ja sisaldavad tavaliselt kahte tuuma [1].
HepG2-rakkude mutatsiooniprofiil
HepG2 rakuliin kannab TERT promootorpiirkonna mutatsiooni C228T, mis esineb ka hepatotsellulaarse kartsinoomi (HCC) ja hepatoblastoomi (HB) puhul. See mutatsioon soodustab immortalisatsiooni, kaitstes vähirakkude telomeere. Lisaks on HepG2 rakkudel looduslik TP53, mis on inimese vähktõve pärssimisel kriitiline geen, kuna see mängib rolli rakutsükli peatamises, apoptoosis ja vananemises. Selle geeni mutatsioonid võivad soodustada rakkude proliferatsiooni.
HepG2 rakud osalevad mitmes signaalitees, sealhulgas rakkude kasvu regulatsiooni häiretes, ellujäämisradades, nagu loote- ja embrüonaalne HB, ning Wnt/β-kateniini signaalitees. Lisaks on sellel rakuliinil iseloomulik CTNNB1-geeni kolmanda eksoni deletsioon, mis on identne epiteel-tüüpi HB-s täheldatuga [2,3].

Ülevaade HepG2-rakuliinist maksarakulise kartsinoomi uurimisel
Inimese hepatomast pärinevad HepG2 rakud on muutunud hindamatuks abivahendiks maksa funktsioonide ja haiguste, sealhulgas hepatotsellulaarse kartsinoomi uurimisel. Need maksarakuliinid annavad ülevaate inimese hepatotsüütide rakulistest reaktsioonidest erinevates katsetingimustes. Lusiferaasi reporterplasmide kasutamine HepG2 rakkudes on osutunud eriti efektiivseks geeniekspressiooni ja rakkude transfektsioonide jälgimisel, mis on olulised ainevahetuse uuringutes, näiteks etanooli mõju uurimisel maksarakkudele.
Viirusnakkuste ja maksahaiguste uuringud HepG2-rakkude abil
Immortaliseeritud maksavähi rakuliinid, nagu HepG2 ja Huh7, on olulised viirusinfektsioonide uurimisel, näidates hepatiit D (HDV) täielikku rakutsükli replikatsiooni ja hepatiit B (HBV) ekspressiooni [5,6]. Samal ajal mängivad HepaRG rakuliinid olulist rolli HBV sisenemismehhanismide selgitamisel [7]. HepG2-rakke kasutatakse ka mitmesuguste inimese maksahaiguste uurimiseks, alates geneetilistest haigustest, nagu progressiivne perekondlik intrahepaatiline kolestaas (PFIC) ja Dubin-Johnsoni sündroom, kuni tsütotoksiliste ja genotoksiliste ainete keskkonnaja toitumisuuringuteni, samuti ravimite sihtmärgistamise ja hepatokartsinogeneesi uurimisel [8,9]. Nende kasutamine ulatub ka bio-kunstliku maksa seadmetega tehtavatesse katsetesse.
HepG2-rakkude interaktsioonid biomaterjalidega koetehnoloogias
HepG2-rakkude interaktsioon erinevate biomaterjalidega on kudede inseneriteaduses otsustava tähtsusega. Sellised tehnikad nagu kolloidproovi tehnika aitavad neid interaktsioone mõista, mõõtes rakkude adhesiivseid omadusi, mis on olulised rakkude eluvõimelisuse kindlaksmääramisel rakkude skelettide ja täpsete maksakoe mudelite arendamiseks.
Rakkude käitumine ja uuendused HepG2-põhistes mudelites
Rakkude käitumise uurimine HepG2-põhistes mudelites on maksa haiguste uurimisel otsustava tähtsusega. Kolmemõõtmeliste sfääriliste rakukultuuride areng on viinud HepG2-rakkude sfääride loomiseni, pakkudes füsioloogiliselt asjakohasemat mudelit, mis peegeldab täpselt normaalsete hepatotsüütide käitumist. Need 3D-mudelid, millel on suurenenud ainevahetusaktiivsus, viitavad HepG2-rakkude potentsiaalile toimida hepatoblastoomi mudelina ning on olulised vähi ravi uurimisel, eriti maksatuumorite simuleerimisel ja uute ravivõtete katsetamisel [10–12].
HepG2 võrdlus ja omadused teiste kasvajarakuliinide seas
HepG2 on üks enim kasutatavaid maksavähi rakuliine, mis on valitud selle laiaulatusliku rakendatavuse tõttu teadusuuringutes umbes 40 kättesaadava maksavähi rakuliini hulgast [13]. Hoolimata teatud tsütokroom P450 ensüümide nõrgast või puuduvast ekspressioonist võrreldes normaalse hepatotsüütidega, on HepG2 metaboolne profiil ajendanud pingutusi rakuliini modifitseerimiseks, et parandada ravimite metabolismi uuringuid [13]. Võrreldes selliste kasvajarakuliinidega nagu MCF7, PC3, 143B ja HEK293, on HepG2 rakkudel unikaalsed aminohapete sisalduse profiilid, mis mõjutavad oluliselt valkude sünteesi ja sekretsiooni, rõhutades nende unikaalseid ainevahetusradusid [14].
HepG2 abil maksahaiguste uurimise tutvustus
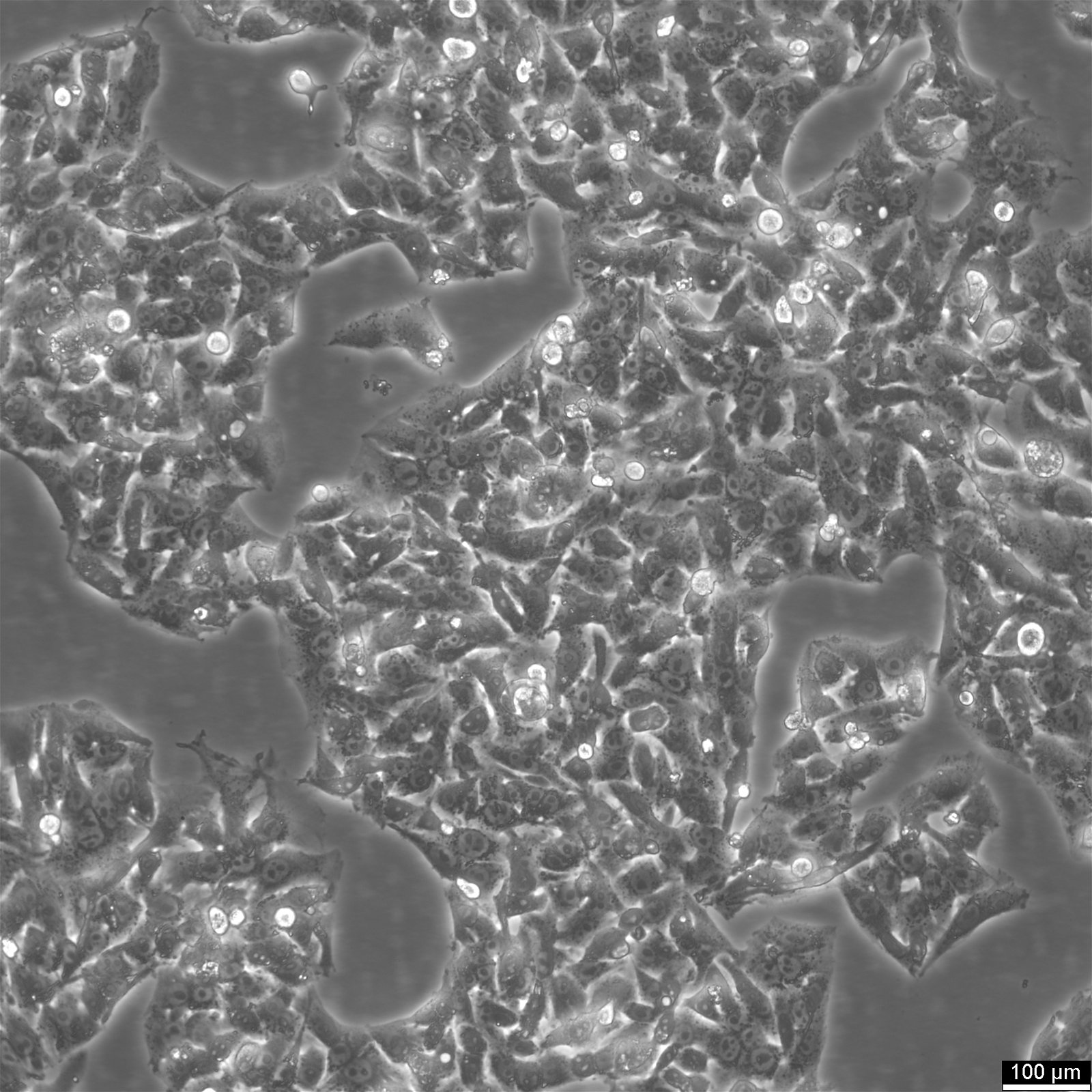
HepG2-rakkude edasikasvatamine
Siin on viis sammu adheseerunud rakkude eemaldamiseks rakukultuurikolvidest Accutase'i abil:
- Eemaldage keskkond rakukultuuri kolvist ja loputage adheseerunud rakud kaltsiumi- ja magneesiumivaba PBS-ga. Kasutage T25 kolvide puhul 3–5 ml PBS-i ja T75 kolvide puhul 5–10 ml PBS-i.
- Lisage Accutase rakukultuuri kolbi, kasutades 1–2 ml T25 ja 2,5 ml T75 kolbi kohta. Veenduge, et Accutase katab kogu rakukihi.
- Inkubeerige kolbi toatemperatuuril 8–10 minutit.
- Resuspendeerige rakud ettevaatlikult 10 ml värske keskkonnaga.
- Tsentrifuugige resuspendeeritud rakke 5 minutit kiirusel 300xg, resuspendeerige need värskes keskkonnas ja jaotage need uutesse kolbidesse, mis sisaldavad värsket keskkonda.
HepG2 rakkude tulevikuväljavaated
Püüdlused HepG2 rakuliini täieliku potentsiaali avamiseks jätkuvad murranguliste edusammudega tsütokroomide ekspressiooni suurendamisel. Teadlased uurivad ka kolmemõõtmeliste sfääriliste rakukultuuride võimalusi, mis pakuvad füsioloogiliselt asjakohasemat süsteemi. Metaboolne aktiivsus, sealhulgas tsütokroomid, on 3D sfäärilistes HepG2 mudelites märkimisväärselt kõrgem kui 2D rakkudes, mis viib meid lähemale normaalseid hepatotsüüte peegeldava mudeli loomisele. Lisaks võib rakupinna valkude ebaõige jaotumise aluseks olevate dünaamiliste protsesside uurimine sillutada teed maksahaiguste paremaks mõistmiseks.
HepG2-rakud: nende rolli ja eripärade mõistmine biomeditsiinilises uurimistöös – KKK
Viited
- Vyas, R.C., Darroudi, F., Natarajan, A.T. Kiirgusest tingitud kromosoomide murdumine ja taasühinemine inimese lümfotsüütide interfase-metafaasi kromosoomides, Mutat Res, 1991; 249(1):29-35.
- Woodfield, S.E., Shi, Y., Patel, R.H., Chen, Z., Shah, A.P., Srivastava, R.K., Whitlock, R.S., Ibarra, A.M., Larson, S.R., Sarabia, S.F., jt. MDM4 inhibeerimine: uus ravistrateegia p53 reaktiveerimiseks hepatoblastoomis. Sci. Rep. 2021, 11, 2967.
- Hussain, S.P., Schwank, J., Staib, F., Wang, X.W., Harris, C.C. TP53-mutatsioonid ja hepatotsellulaarne kartsinoom: ülevaade maksavähi etioloogiast ja patogeneesist. Oncogene 2004.
- Schicht, G., Seidemann, L., Haensel, R., Seehofer, D., Damm, G. Hepatooma rakuliinide HepG2 ja Huh7 kasutatavuse kriitiline uurimine resekteeritava hepatotsellulaarse kartsinoomi metaboolse esindatuse mudelitena. Cancers 2022, 14(17), 4227.
- Verrier, E.R., Colpitts, C.C., Schuster, C., Zeisel, M.B., Baumert, T.F. Rakukultuurimudelid B- ja D-hepatiidi viirusnakkuse uurimiseks. Viruses 2016, 8, 261.
- Verrier, E.R., Colpitts, C.C., Bach, C., Heydmann, L., Weiss, A., Renaud, M., Durand, S.C., Habersetzer, F., Durantel, D., AbouJaoudé, G. jt. Sihtotstarbeline funktsionaalne RNA-interferentsi sõelumine paljastab glükopaan 5 kui B- ja D-hepatiidi viiruste sisenemisfaktori. Hepatology 2016, 63, 35–48.
- Gripon, P., Rumin, S., Urban, S., Le Seyec, J., Glaise, D., Cannie, I., Guyomard, C., Lucas, J., Trepo, C., Guguen-Guillouzo, C. Inimese hepatoma rakuliini nakatumine B-hepatiidi viirusega. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660.
- Mersch-Sundermann, V., Knasmüller, S., Wu, X.J., Darroudi, F., Kassie, F. Inimese maksa rakuliini kasutamine tsütoprotektiivsete, antigenotoksiliste ja kogenotoksiliste ainete avastamiseks. Toxicology. 2004; 198(1–3): 329–340.
- Fanelli, A. HepG2 (maksa hepatotsellulaarne kartsinoom): rakukultuur. HepG2. Vaadatud 3. detsembril 2017.
- Xuan, J., Chen, S., Ning, B., Tolleson, W.H., Guo, L. Tsütokroom P450-sid ekspresseerivate HepG2-põhiste rakkude arendamine ainevahetusega seotud ravimite poolt indutseeritud maksatoksilisuse hindamiseks. Physiol. Behav. 2017, 176, 139–148.
- Ooka, M., Lynch, C., Xia, M. In vitro ainevahetuse aktiveerimise rakendamine suure läbilaskevõimega sõelumisel. Int. J. Mol. Sci. 2020, 21, 8182.
- Huang, L., Coughtrie, M.W.H., Hsu, H. Dehüdroepiandrosteroonisulfotransferaasi geeni allaregulatsioon inimese hepatotsellulaarses kartsinoomis. Mol. Cell. Endocrinol.
- Zhu, Z., Hao, X., Yan, M., jt. Vähirakkude tüvi- ja eellastrakud on maksarakulise kartsinoomi puhul CD133 + CD44 + populatsioonis väga rikkalikult esindatud. Int J Cancer. 2010; 126:2067-2078.
- Arbus, C., Benyamina, A., Llorca, P.-M., Baylé, F., Bromet, N., Massiere, F., Garay, R.P., Hameg, A. Tsüamemasiini metabolismis osalevate inimese tsütokroom P450 ensüümide iseloomustus. Eur J Pharm Sci. 2007 dets;32(4-5):357-66.