
, L-alanyl-L-glutaminlösning")
Volym: 100 ml
Förvaring: ≤ –15 °C
Sterilitet: Sterilfiltrerad
Stabil glutaminlösning (L-alanyl-L-glutamin, 200 mM) är en mycket stabil dipeptidform av L-glutamin som är utformad som en direkt ersättning för konventionell L-glutamin i cellodlingsmedier. L-glutamin är en essentiell aminosyra och en viktig energikälla för odlade celler, som spelar en avgörande roll för celltillväxt, metabolism och proteinsyntes.
Användning och fördelar
I standardvätskemedia bryts L-glutamin ned relativt snabbt vid 37 °C, vilket leder till bildandet av toxiska biprodukter såsom ammoniumjoner som kan påverka cellernas livskraft och experimentresultaten negativt. Stabil glutamin övervinner denna begränsning genom att tillhandahålla en icke-nedbrytbar dipeptidform som förblir intakt under odlingsförhållanden.
Cellerna klyver enzymatiskt dipeptidbindningen för att frigöra L-glutamin efter behov, vilket säkerställer en kontinuerlig färsk tillförsel samtidigt som ackumulering av skadliga avfallsprodukter förhindras. Detta gör lösningen särskilt fördelaktig för långvariga cellodlingar och odlingssystem med hög densitet.
Formulering och användning
L-Alanyl-L-Glutamin-lösningen bereds i vatten av cellodlingskvalitet i en koncentration på 200 mM och är sterilfiltrerad för kontamineringskänsliga tillämpningar. Den kan spädas direkt i komplett medium enligt experimentella krav. Förvara vid ≤ –15 °C och undvik upprepade frys-tina-cykler för att bibehålla produktens stabilitet.
Endast för forskningsändamål. Får inte användas i diagnostiska eller terapeutiska procedurer. Får inte användas på människor eller djur.
")
Volym: 100 ml
Förvaring: +2 °C till +8 °C
Sterilitet: Sterilfiltrerad
HEPES-buffertlösning (1 M), även känd som N-2-hydroxietylpiperazin-N-2-etansulfonsyra, är ett zwitterjoniskt organiskt buffertmedel som ofta används i cellodlingsmedier. Det är utformat för att upprätthålla stabila pH-förhållanden inom det fysiologiska intervallet 6,7 till 8,6, vilket stödjer optimal cellfunktion vid in vitro-tillämpningar.
Användning och fördelar
HEPES ger tillförlitlig buffertkapacitet i cellodlingssystem, särskilt när celler hanteras utanför en CO₂-inkubator. Tillsatsen av 10–25 mM HEPES till odlingsmediet ger förbättrad pH-stabilitet under längre hanteringsperioder, vilket bidrar till att upprätthålla konsekventa experimentella förhållanden.
Denna buffert är membranimpermeabel, har minimal inverkan på biokemiska reaktioner och uppvisar stark kemisk och enzymatisk stabilitet. Dessa egenskaper gör den lämplig för ett brett spektrum av cellodlings
- och biokemiska tillämpningar.
Formulering och användning
Lösningen levereras som ett 1 M koncentrat berett i vatten av cellodlingskvalitet och är steriliserat för användning i kontamineringskänsliga miljöer. Den kan spädas till önskad arbets koncentration beroende på tillämpningskraven. Förvara vid +2 °C till +8 °C och hantera under aseptiska förhållanden för att bevara produktens integritet.
Endast för forskningsändamål. Får inte användas i diagnostiska eller terapeutiska procedurer. Får inte användas på människor eller djur.
, 250 g/l")
Volym: 50 ml
Förvaring: +2 °C till +8 °C
Sterilitet: Sterilfiltrerad
D-(+)-glukoslösning (dextroslösning) är ett sterilt, färdigt att använda tillskott som innehåller det naturligt förekommande sockret D-(+)-glukos, en central komponent i cellmetabolismen. Glukos är involverat i viktiga biologiska processer såsom energiproduktion, glykosylering och bildandet av glykaner som bidrar till cellernas struktur och funktion.
Användning och fördelar
Denna glukoslösning används ofta som tillskott i cellodlingsmedier och i många cellulära och molekylärbiologiska tillämpningar. Som primär koldioxid
- och energikälla stöder glukos celltillväxt, proliferation och metabolisk aktivitet. Dess inblandning i biosyntetiska vägar gör den också avgörande för att upprätthålla normal cellfysiologi och experimentell konsistens.
Formulering och användning
Lösningen levereras i en hög koncentration på 250 g/l glukos, vilket möjliggör flexibel utspädning i odlingsmedier enligt experimentella krav. Den är sterilfiltrerad för att säkerställa lämplighet för kontamineringskänsliga tillämpningar. Förvara vid +2 °C till +8 °C och hantera aseptiskt för att bibehålla produktens kvalitet och stabilitet.
Endast för forskningsändamål. Får inte användas i diagnostiska eller terapeutiska procedurer. Får inte användas på människor eller djur.
, lösning 100x")
Volym: 10 ml
Förvaring: +2 °C till +8 °C
Sterilitet: Sterilfiltrerad
Insulin-Transferrin-Selenium (ITS) Solution (100x) är ett kemiskt definierat tillskott avsett för ett brett spektrum av cellodlingsapplikationer. Det används oftast som tillsats i basala cellodlingsmedier för att stödja celltillväxt under förhållanden med reducerat serum eller utan serum.
Användning och fördelar
Vårt ITS-tillskott tillhandahåller väsentliga komponenter som krävs för optimal prestanda hos serumfria medier. Genom att komplettera konventionella näringsmedier med ITS kan behovet av fetalt bovint serum (FBS) för rutinunderhåll av många cellinjer minskas avsevärt. Detta bidrar till att minimera variabiliteten förknippad med serumanvändning samtidigt som en jämn celltillväxt och livskraft upprätthålls.
Insulin stöder cellernas upptag och metabolism av viktiga näringsämnen, transferrin underlättar järntransporten och selen bidrar till antioxidantförsvaret och den enzymatiska aktiviteten. Tillsammans främjar dessa komponenter en balanserad cellmetabolism och förbättrad reproducerbarhet i definierade odlingssystem.
Sammansättning och användning
Insulin-Transferrin-Selenium (ITS) levereras som en 100-faldigt koncentrerad lösning i Earle's Balanced Salt Solution (EBSS) utan fenolrött. För standardtillämpningar, späd 1:100 i lämpligt basmedium för att uppnå den rekommenderade arbetskoncentrationen. Förvara vid +2 °C till +8 °C och hantera under aseptiska förhållanden för att bibehålla produktens stabilitet och sterilitet.
Endast för forskningsändamål. Får inte användas i diagnostiska eller terapeutiska procedurer. Får inte användas på människor eller djur.
")
Volym: 5 ml
Förvaring: +2 °C till +8 °C
Sterilitet: Sterilfiltrerad
Insulin Human Recombinant Solution är ett kemiskt definierat tillskott som vanligtvis används för odling av däggdjurscellinjer, inklusive celler från kinesiska hamstrars äggstockar (CHO-celler). Denna lösning av cellodlingskvalitet innehåller rekombinant humant insulin uttryckt i Saccharomyces cerevisiae, vilket garanterar hög renhet och jämn prestanda i forskningsapplikationer.
Användning och fördelar
Insulin tillsätts rutinmässigt till serumfria och kemiskt definierade medier för att främja celltillväxt och produktivitet. Som ett viktigt reglerande hormon stöder insulin cellernas upptag, utnyttjande och lagring av glukos, aminosyror och fettsyror. Det hämmar också nedbrytningen av glykogen, proteiner och lipider, vilket bidrar till förbättrad cellviabilitet och metabolisk stabilitet i odlingssystem. Den kemiskt definierade formuleringen stöder reproducerbarhet och minimerar variabiliteten i känsliga cellodlingsarbetsflöden.
Biologiska egenskaper och användning
Insulin är ett tvåkedjigt polypeptidhormon som produceras naturligt av β-cellerna i bukspottkörtelns öar. Det har en molekylvikt på cirka 5 800 Da. α
- och β-kedjorna är bundna av två disulfidbindningar mellan kedjorna, och α-kedjan innehåller en disulfidbindning inom kedjan. För cellodlingsapplikationer ska lösningen hanteras under aseptiska förhållanden och förvaras vid +2 °C till +8 °C för att bibehålla stabilitet och prestanda.
Endast för forskningsändamål. Får inte användas i diagnostiska eller terapeutiska procedurer. Får inte användas på människor eller djur.
")
Volym: 100 ml
Förvaring: +2 °C till +8 °C
Sterilitet: Sterilfiltrerad
Natriumpyruvatlösning (100 mM) är ett sterilt, färdigt att använda tillskott som är utformat för att ge en extra, lättillgänglig energikälla för cellodlingsmedier. Natriumpyruvat spelar en viktig roll i cellernas energimetabolism och stöder tillväxten av metaboliskt aktiva och snabbt prolifererande celler, såsom tumörceller. Tillskott kan förbättra cellernas livskraft och bidra till att upprätthålla metabolisk stabilitet i odlingssystem.
Användning och fördelar
Denna lösning används ofta i rutinmässig cellodling för att berika medier med pyruvat och främja optimala tillväxtförhållanden. Den stöder ATP-produktion, kan bidra till att minska oxidativ stress och bidrar till förbättrad metabolisk prestanda hos odlade celler. Produkten tillverkas i vatten av cellodlingskvalitet och är sterilt filtrerad, vilket garanterar jämn kvalitet och reproducerbarhet i forskningsarbetsflöden.
Användning och kompatibilitet
Den rekommenderade slutkoncentrationen för de flesta cellodlingsapplikationer är 1 mM natriumpyruvat, vilket uppnås genom att späda 100 mM stamlösningen 1:100 i komplett odlingsmedium. Lösningen är kompatibel med ett brett spektrum av basala medier och däggdjurscellinjer. Förvara vid +2 °C till +8 °C och skydda mot kontaminering för att bibehålla produktens stabilitet.
Endast för forskningsändamål. Får inte användas i diagnostiska eller terapeutiska procedurer. Får inte användas på människor eller djur.
")
Volym: 100 ml Förvaring: ≤-15°C Sterilitet: Sterilt filtrerad
Antibiotic/Antimycotic Solution (100x) är ett sterilt, bruksfärdigt koncentrat som är utformat för att minska riskerna för mikrobiell kontaminering vid cellodling och relaterade laboratorieapplikationer. Denna 100x-lösning innehåller en väletablerad kombination av penicillin, streptomycin och amfotericin B, vilket ger en bredspektrig antimikrobiell aktivitet mot grampositiva och gramnegativa bakterier, jästsvampar och filamentösa svampar. Formuleringen är lämplig för användning i eukaryota cellkulturer, bakteriemedier och andra kontaminationskänsliga system, vilket stöder en ren och konsekvent laboratorieverksamhet.
Tillämpning och fördelar Denna lösning är optimerad för rutinmässiga forskningsprotokoll och används ofta för att upprätthålla aseptiska förhållanden i arbetsflöden för cellodling. Den ger tillförlitlig prestanda i kontamineringskänsliga miljöer och hjälper forskare att minska risken för mikrobiell överväxt utan att äventyra cellernas hälsa eller experimentens reproducerbarhet. Den sterilfiltrerade formuleringen eliminerar behovet av ytterligare solubiliseringssteg, vilket ger en strömlinjeformad medieberedning och minskar variabiliteten i de dagliga laboratorieprocedurerna.
Användning och kompatibilitet För att uppnå standardiserade arbetskoncentrationer, späd lösningen 1:100 i ditt kompletta odlingsmedium. Produkten är kompatibel med ett brett spektrum av mammaliecellinjer och basala medier. Med konsekvent lagertillgänglighet kan forskare dra nytta av pålitlig leveranskontinuitet och förenklad logistikplanering. Lösningen ska förvaras vid ≤ -15 °C och skyddas från upprepade frys
- och upptiningscykler för att bibehålla stabiliteten. Endast för forskningsändamål. Ej för användning i diagnostiska eller terapeutiska procedurer. Ej för användning på människor eller djur.
")
Volym: 100 ml Förvaring: +2°C till +8°C Sterilitet: Sterilt filtrerad
MEM Non-Essential Amino Acids (100x) är ett sterilt tillskott som är utformat för att förbättra celltillväxt och livskraft i cellodlingssystem för däggdjur. Formuleringen motsvarar ett 100x koncentrat av de icke-essentiella aminosyror som finns i standard Minimum Essential Medium (MEM), vilket möjliggör direkt tillskott av basala medier med minimal beredning.
Tillämpning och fördelar Detta tillskott ger en extra aminosyrapool för snabbt prolifererande celler eller för cellinjer som har förlorat förmågan att syntetisera icke-essentiella aminosyror de novo. Genom att lindra den metaboliska bördan av biosyntesen bidrar det till förbättrad tillväxtkinetik, förlängd livsduglighet och större experimentell konsekvens
- särskilt i näringskänsliga kulturer eller kulturer med hög densitet.
Sammansättning och användning Lösningen innehåller glycin, L-alanin, L-asparagin, L-asparaginsyra, L-glutaminsyra, L-prolin och L-serin. Den är kompatibel med MEM och de flesta andra standardmedier. För användning, späd 1:100 i det slutliga odlingsmediet. Denna produkt är sterilfiltrerad och klar att använda utan ytterligare hanteringssteg. Endast för forskningsändamål. Ej för användning i diagnostiska eller terapeutiska procedurer. Ej för användning på människor eller djur.

- Ett skonsamt alternativ till Trypsin
Accutase är en cellavskiljningslösning som revolutionerar cellodlingsindustrin. Det är en blandning av proteolytiska och kollagenolytiska enzymer som efterliknar effekten av trypsin och kollagenas. Till skillnad från trypsin innehåller Accutase inga däggdjurs
- eller bakteriekomponenter och är mycket skonsammare mot cellerna, vilket gör det till en idealisk lösning för rutinmässig avskiljning av celler från vanliga plastbehållare för vävnadsodling och plastbehållare med adhesionsbeläggning. I det här blogginlägget går vi igenom fördelarna och användningsområdena för Accutase och hur det förändrar spelreglerna för cellodling.
Fördelar med Accutase
Accutase har flera fördelar jämfört med traditionella trypsinlösningar. För det första kan det användas när det behövs en skonsam och effektiv avskiljning av en adherent cellinje, vilket gör det till en direkt ersättning för trypsin. För det andra fungerar Accutase mycket bra på embryonala och neuronala stamceller, och det har visat sig att dessa cellers viabilitet bibehålls efter passering. För det tredje bevarar Accutase de flesta epitoper för efterföljande flödescytometrianalys, vilket gör den idealisk för analys av cellytemarkörer.
Dessutom behöver Accutase inte neutraliseras vid passering av adherenta celler. Genom att tillsätta mer media efter att cellerna har delats späds Accutase ut så att det inte längre kan lossa cellerna. Detta eliminerar behovet av ett inaktiveringssteg och sparar tid för cellodlingsteknikerna. Slutligen behöver Accutase inte alikeras, och en flaska är stabil i kylskåpet i 2 månader.
Tillämpningar av Accutase
Accutase är en direkt ersättning för trypsinlösning och kan användas för passering av cellinjer. Dessutom fungerar Accutase bra när celler lossas för analys av många cellytmarkörer med hjälp av flödescytometri och för cellsortering. Andra nedströmsapplikationer för Accutase-behandling inkluderar analys av cellytmarkörer, virustillväxtanalys, cellproliferation, tumörcellsmigrationsanalyser, rutinmässig cellpassage, produktionsuppskalning (bioreaktor) och flödescytometri.
Sammansättning av Accutase
Accutase innehåller inga däggdjurs
- eller bakteriekomponenter och är en naturlig enzymblandning med proteolytisk och kollagenolytisk enzymaktivitet. Den är formulerad i en mycket lägre koncentration än trypsin och kollagenas, vilket gör den mindre toxisk och skonsammare, men lika effektiv.
Effektiviteten hos Accutase
Accutase har visat sig vara effektivt när det gäller att avlägsna primära celler och stamceller och bibehålla hög cellviabilitet jämfört med enzymer av animaliskt ursprung, t.ex. trypsin. 100% av cellerna återvinns efter 10 minuter, och det är ingen fara att låta cellerna ligga i Accutase i upp till 45 minuter, tack vare Accutases autodigestion.
Sammanfattningsvis
Sammanfattningsvis är Accutase en kraftfull lösning som håller på att förändra spelreglerna för cellodling. Med sin skonsamma natur, effektivitet och mångsidighet är Accutase det perfekta alternativet till trypsin. Om du letar efter en tillförlitlig och effektiv lösning för cellavskiljning är Accutase lösningen för dig.
, w: 2 mM L-Glutamin, w: 2,2 g/L NaHCO3, w: EBSS")
Vad finns i EMEM?
EMEM är en modifierad version av Eagle's minimum essential medium, som innehåller Earle's Balanced Salt Solution, icke-essentiella aminosyror, L-glutamin, natriumpyruvat och natriumbikarbonat. Det är viktigt att notera att denna nivå av natriumbikarbonat är avsedd för användning i 5% CO2 i luften. För att bibehålla dess effektivitet rekommenderas att mediet förvaras mörkt vid 2°C till 8°C när det inte används.
Vad används EMEM till?
Eagle's minimal essential medium (EMEM) är ett cellodlingsmedium som kan underhålla celler i vävnadsodling. Mediet innehåller högre koncentrationer av aminosyror, vilket möjliggör en mer exakt approximation av proteinsammansättningen i odlade däggdjursceller. EMEM kan användas för att odla olika celler, bland annat fibroblaster, levercancerceller från människa (HepG2) och astrocytceller från människa (PDA) som härrör från fosterhjärnans progenitor. Det används vanligen i närvaro av fetalt bovint serum (FBS), kalv
- eller hästserum.
Hur skiljer sig EMEM från andra cellodlingsmedier?
EMEM och Dulbeccos modifierade Eagle's medium (DMEM) har vissa likheter, men de skiljer sig också åt. Båda medierna saknar protein och innehåller de aminosyror, salter, glukos och vitaminer som krävs för att förse en cell med energi och upprätthålla den i vävnadskultur. DMEM-formuleringen är dock modifierad så att den innehåller upp till fyra gånger mer vitaminer och aminosyror och två till fyra gånger mer glukos än EMEM. Det är värt att notera att EMEM också skiljer sig från den ursprungliga MEM-formuleringen.
Kvalitetskontroll
Sterilt filtrerad
Förvaring och hållbarhetstid
Förvaras i +2°C till +8°C, skyddat från ljus.
Efter öppning, förvara vid 4°C och använd inom 6-8 veckor.
Förhållanden vid leverans
Omgivande temperatur
Underhåll
Förvaras i kylskåp vid +2°C till +8°C i mörker. Undvik frysning och frekvent uppvärmning till +37°C, eftersom det försämrar produktkvaliteten.
Värm inte upp mediet över 37°C och använd inte okontrollerade värmekällor som t.ex. mikrovågsugnar.
Om endast en del av mediet ska användas, ta ut den mängd som behövs och värm det till rumstemperatur före användning.
Sammansättning
Kategori
Komponenter
Koncentration (mg/L)
Aminosyror
L-arginin HCl
126.00
L-kystin 2 HCl
31.30
L-Glutamin
292.00
L-Histidin HClH2O
42.00
L-Isoleucin
52.00
L-Leucin
52.00
L-Lysin HCl
72.50
L-metionin
15.00
L-fenylalanin
32.00
L-Treonin
48.00
L-tryptofan
10.00
L-tyrosin 2 Na 2H2O
51.90
L-Valin
46.00
Vitaminer
Kolinklorid
1.00
Vitaminer
D-kalciumpantotenat
1.00
Folsyra
1.00
myo-Inositol
2.00
Nikotinamid
1.00
Pyridoxal HCl
1.00
Riboflavin
0.10
Tiamin HCl
1.00
Oorganiska salter
CaCl2 2H2O
265.00
Oorganiska salter
KCl
400.00
MgSO4
97.67
NaCl
6800.00
NaHCO3
2200.00
NaH2PO4
122.00
Övriga komponenter
D-Glukos
1000.00
Övriga komponenter
Fenolrött natriumsalt
11.00

Viktiga egenskaper hos Freeze Medium CM-1 inkluderar:
Bred kompatibilitet: Effektivt för ett stort antal olika celltyper, inklusive primära celler, stamceller och etablerade cellinjer.
Hög viabilitet: Optimerat för att maximera cellåtervinning och viabilitet efter upptining, vilket säkerställer tillförlitliga experimentella resultat.
Redo att använda: Bekvämt förberedd och steriliserad för omedelbar användning, vilket minskar förberedelsetiden och risken för kontaminering.
Förbättrad stabilitet: Upprätthåller konsekvent prestanda under standardförhållanden för kryokonservering, vilket säkerställer reproducerbara resultat.
Lång hållbarhetstid: CM-1 är ett serumhaltigt kryopreserveringsmedium som är färdigt att använda och kan förvaras i kylskåp i upp till ett år.
Användning av CM-1 för frysning av celler
Följ dessa steg för att använda CM-1 för frysning av både adherenta celler och suspenderade celler
För adherenta celler, tvätta och separera dem från odlingssubstratet. För suspenderade celler, fortsätt direkt till nästa steg.
Räkna cellerna för att säkerställa att de har rätt koncentration.
Centrifugera cellerna för att pelletera dem och resuspendera sedan i CM-1-frysmedium.
Överför de resuspenderade cellerna till kryovialer.
Använd en metod med långsam frysning innan cellerna överförs till långtidsförvaring
Metod
Beskrivning av metoden
Steg för steg
❄️
Manuell frysning
En steg-för-steg-metod som innebär gradvis temperatursänkning för att säkerställa cellernas livskraft
1️⃣ Placera cellerna i frysmedium i en 4°C frys i 40 minuter.
2️⃣ Överför till en frys på -80°C i 24 timmar.
3️⃣ Förvara cellerna i flytande kväve för långtidsförvaring
❄️
Användning av Mr Frosty
En praktisk anordning som möjliggör kontrollerade fryshastigheter utan elektrisk ström
1️⃣ Förbered celler i kryovialer med frysmedium.
2️⃣ Placera kryovialerna i Mr Frosty-behållaren.
3️⃣ Förvara vid -80°C i 24 timmar innan du överför till flytande kväve
❄️
Frys med kontrollerad hastighet
En högprecisionsfrys från Thermo Fisher eller andra tillverkare som är utformad för kontrollerad temperatursänkning
1️⃣ Programmera enheten för att gradvis sänka temperaturen.
2️⃣ Placera de förberedda cellerna i frysen.
3️⃣ Efter fryscykeln, överför cellerna till flytande kväve
Förvara kryovialerna vid temperaturer under -130°C eller i flytande kväve för långtidsförvaring.
Ingredienser
Innehåller FBS, DMSO, glukos, salter
Buffringskapacitet: pH = 7,2 till 7,6
Cytions Freeze Medium CM-1 erbjuder en tillförlitlig lösning för kryokonservering, vilket säkerställer hög cellviabilitet och funktionalitet efter upptining för ett brett spektrum av forskningsapplikationer.

- och kycklingleverceller, särskilt under förhållanden med reducerat serum.
Ham's F-12K (Kaighn's) Medium är noggrant formulerat för att optimera cellodlingsförhållandena. Det har en berikad sammansättning som ger förhöjda nivåer av essentiella komponenter som aminosyror och natriumpyruvat, samt ytterligare element som putrescin, tymidin, hypoxantin och zink. Dessa tillsatser gör det möjligt för forskare att komplettera mediet med minimalt serum eller definierade komponenter för specifika celltyper, vilket underlättar exakta experimentella förhållanden.
Ham's F-12K (Kaighn's) Medium innehåller inte proteiner eller tillväxtfaktorer. Därför är det ofta nödvändigt att komplettera med tillväxtfaktorer och Fetal Bovine Serum (FBS), vilket gör det möjligt för forskare att skräddarsy mediet efter kraven för sina specifika cellinjer. För optimal prestanda måste koncentrationen av FBS noggrant optimeras för varje cellinje, så att optimal tillväxt och funktionalitet säkerställs.
För att upprätthålla ett fysiologiskt pH använder Ham's F-12K (Kaighn's) Medium ett natriumbikarbonatbuffert-system (2,5 g/L), vilket kräver en kontrollerad 5-10% CO2-miljö under odlingen. Detta säkerställer att mediets pH-värde ligger inom det idealiska intervallet för celltillväxt och livskraft
Kvalitetskontroll
pH = 7,2 +/
- 0,02 vid 20-25°C.
Varje parti har testats för sterilitet och frånvaro av mykoplasma och bakterier.
Förvaring
Förvaras i kylskåp vid +2°C till +8°C i mörker. Frysning och uppvärmning upp till +37°C minimerar produktens kvalitet.
Värm inte upp mediet till mer än 37°C och använd inte okontrollerbara värmekällor (t.ex. mikrovågsugnar).
Om endast en del av mediet ska användas, ta bort denna mängd från flaskan och värm upp den i rumstemperatur.
Hållbarheten för alla medier utom basmediet är 8 veckor från tillverkningsdatum.
Sammansättning
Komponenter
mg/L
Oorganiska salter
Kalciumklorid x 2H2O
135,24
Koppar(II)sulfat x 5H2O
0,00
Järn(II)sulfat x 7H2O
0,83
Magnesiumklorid x 6H2O
105,72
Magnesiumsulfat x 7H2O
394,49
Kaliumklorid
283,29
Kaliumdivätefosfat
58,52
Natriumklorid
7597,20
di-Natriumvätefosfatvattenfri
115,02
Zinksulfat x 7H2O
0,14
Övriga komponenter
D(+)-Glukos vattenfri
1260,00
Hypoxanthin
4,08
DL-α-lipoinsyra
0,21
Fenol röd
3,00
Putrescin x 2HCl
0,32
Natrium pyruvat
220,00
NaHCO3
2500,00
Tymidin
0,73
Aminosyror
L-Alanin
17,82
L-arginin x HCl
421,40
L-asparagin x H2O
30,02
L-Asparaginsyra
26,62
L-Cystein x HCl x H2O
70,24
L-glutamin
292,20
L-glutaminsyra
29,42
Glycin
15,01
L-Histidin x HCl x H2O
41,92
L-Isoleucin
7,87
L-Leucin
26,24
L-Lysin x HCl
73,04
L-metionin
8,95
L-fenylalanin
9,91
L-prolin
69,06
L-Serin
21,02
L-Threonin
23,82
L-tryptofan
4,08
L-tyrosin
10,87
L-Valin
23,42
Vitaminer
D(+)-Biotin
0,07
D-kalciumpantotenat
0,48
Kolinklorid
13,96
Folsyra
1,32
myo-Inositol
18,02
Nikotinamid
0,04
Pyridoxin x HCl
0,06
Riboflavin
0,04
Tiamin x HCl
0,34
Vitamin B12
1,36

Fosfatbuffrad saltlösning (PBS) är en buffertlösning som används i stor utsträckning inom biologisk och kemisk forskning. Den spelar en avgörande roll för att upprätthålla pH-balansen och osmolariteten under olika experimentella procedurer, inklusive vävnadsbearbetning och cellodling. Vår PBS-lösning är noggrant formulerad med ingredienser med hög renhet för att säkerställa stabilitet och tillförlitlighet i varje experiment. Osmolariteten och jonkoncentrationerna i vår PBS efterliknar i hög grad de som finns i människokroppen, vilket gör den isotonisk och giftfri för de flesta celler.
Sammansättning av vår PBS-lösning
Vår PBS-lösning är en pH-justerad blandning av fosfatbuffertar och saltlösningar av ultrapur kvalitet. Vid en 1X arbetskoncentration innehåller den:
8000 mg/L natriumklorid (NaCl)
200 mg/L kaliumklorid (KCl)
1150 mg/L Natriumfosfat dibasisk vattenfri (Na2HPO4)
200 mg/L Kaliumfosfat monobasiskt vattenfritt (KH2PO4)
Denna sammansättning säkerställer en optimal pH
- och jonbalans, lämplig för ett brett spektrum av biologiska tillämpningar.
Tillämpningar av vår PBS-lösning
Vår PBS-lösning är idealisk för olika tillämpningar inom biologisk forskning. Dess isotoniska och giftfria egenskaper gör den lämplig för utspädning av substanser och sköljning av cellbehållare. PBS-lösningar som innehåller EDTA är effektiva för att frigöra fastsittande och klumpade celler. Tvåvärda metaller som zink bör dock inte tillsättas i PBS, eftersom det kan orsaka utfällning. I sådana fall rekommenderas Good's buffertar. Dessutom är vår PBS-lösning ett godtagbart alternativ till viralt transportmedium för transport och förvaring av RNA-virus, inklusive SARS-CoV-2.
Kvalitetskontroll
Sterilt filtrerad
Förvaring och hållbarhetstid
Förvaras vid +2°C till +25°C, skyddat från ljus.
Efter öppnandet, förvara vid 2°C till 25°C och använd inom 24 månader.
Förhållanden vid leverans
Omgivande temperatur
Underhåll
Förvaras i kylskåp vid +2°C till +8°C i mörker. Undvik frysning och frekvent uppvärmning till +37°C, eftersom det försämrar produktkvaliteten.
Värm inte upp mediet över 37°C och använd inte okontrollerade värmekällor som t.ex. mikrovågsugnar.
Om endast en del av mediet ska användas, ta ut den mängd som behövs och värm det till rumstemperatur före användning.
Sammansättning
Kategori
Komponenter
Koncentration (mg/L)
Salter
Kaliumklorid
200
Kaliumfosfat monobasiskt vattenfritt
200
Natriumklorid
8000
Natriumfosfat dibasisk vattenfri
1150

RPMI 1640 Medium utformades ursprungligen för att stödja tillväxten av mänskliga leukemiceller i både suspension och monolagerkulturer, men har utvecklats genom modifieringar av forskare och kommersiella leverantörer för att bli lämpligt för ett brett spektrum av däggdjursceller. Det är exceptionellt kompatibelt med cellinjer som HeLa, Jurkat, MCF-7, PC12, PBMC, astrocyter och karcinom.
RPMI 1640 Medium skiljer sig från andra cellodlingsmedier på grund av sin unika sammansättning. Det innehåller en betydande mängd fosfat, aminosyror och vitaminer. Framför allt innehåller det biotin, vitamin B12 och PABA, som saknas i Eagle's Minimal Essential Medium eller Dulbecco's Modified Eagle Medium. Dessutom uppvisar RPMI 1640 Medium signifikant förhöjda koncentrationer av vitaminerna inositol och kolin. Det innehåller dock inte proteiner, lipider eller tillväxtfaktorer. Följaktligen krävs ofta tillsats av 10% fetalt bovint serum (FBS) för att ge optimala förutsättningar för celltillväxt.
Buffringssystemet i RPMI 1640 Medium är beroende av natriumbikarbonat och kräver en 5-10% CO2-miljö för att upprätthålla ett fysiologiskt lämpligt pH. Inkluderingen av reduktionsmedlet glutation skiljer detta medium ytterligare från andra.
Kvalitetskontroll
Sterilt filtrerad
Förvaring och hållbarhetstid
Förvaras vid +2°C till +8°C, skyddat från ljus.
Efter öppnandet, förvara vid 4°C och använd inom 6-8 veckor.
Förhållanden vid leverans
Omgivande temperatur
Underhåll
Förvaras i kylskåp vid +2°C till +8°C i mörker. Undvik frysning och frekvent uppvärmning till +37°C, eftersom det försämrar produktkvaliteten.
Värm inte upp mediet över 37°C och använd inte okontrollerade värmekällor som t.ex. mikrovågsugnar.
Om endast en del av mediet ska användas, ta ut den mängd som behövs och värm det till rumstemperatur före användning.
Sammansättning
Kategori
Komponenter
Koncentration (mg/L)
Aminosyror
Glycin
10.00
L-Alanyl-L-Glutamin
434.40
L-arginin
200.00
L-AsparaginH2O
56.82
L-Asparaginsyra
20.00
L-kystin 2HCl
65.20
L-glutaminsyra
20.00
L-Histidin HClH2O
20.27
L-hydroxi-L-prolin
20.00
L-Isoleucin
50.00
L-Leucin
50.00
L-Lysin HCl
40.00
L-metionin
15.00
L-fenylalanin
15.00
L-prolin
20.00
L-Serin
30.00
L-Threonin
20.00
L-Tryptofan
5.00
L-tyrosin 2Na 2H2O
28.83
L-Valin
20.00
Vitaminer
p-Amino bensoesyra
1.00
D-biotin
0.20
Kolinklorid
3.00
D-kalciumpantotenat
0.25
Folsyra
1.00
myo-Inositol
35.00
Nikotinamid
1.00
Pyridoxin HCl
1.00
Riboflavin
0.20
Tiamin HCl
1.00
Vitamin B12
0.005
Oorganiska salter
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Övriga komponenter
D-Glukos
2000.00
L-glutation reducerad
1.00
Fenolrött natriumsalt
5.30
, w: 3,1 g/L glukos, w: 2,5 mM L-glutamin, w: 15 mM HEPES, w: 0,5 mM natriumpyruvat, w: 1,2 g/L NaHCO3")
Denna unika formulering kombinerar Dulbeccos Modified Eagle Medium (DMEM) och Ham's F-12 (Ham's Nutrient Mixture F-12) i ett exakt förhållande 1:1. Tillsatsen av L-glutamin förbättrar dess sammansättning ytterligare.
DMEM, som härrör från Eagle's Minimal Essential Medium (EMEM), erbjuder en ökad koncentration av aminosyror och vitaminer jämfört med sin föregångare. Ham's F-12 är däremot baserat på Ham's F-10 medium, vilket ger en kompletterande uppsättning essentiella komponenter.
För att stödja optimal celltillväxt är det vanligt att DMEM:Ham's F12 kompletteras med FBS i en typisk koncentration på 5-10%. Detta tillskott är nödvändigt eftersom mediet saknar tillväxthormoner, lipider och proteiner som är avgörande för cellutvecklingen.
DMEM:Ham's F12 innehåller ett pH-buffert-system och kompletteras ofta med fenolrött, en pH-indikator. Odlade celler i DMEM:Ham's F12, eller i något annat medium som använder bikarbonatbuffertsystemet, kräver en kontrollerad CO2-miljö på 5-10% för att upprätthålla lämpliga pH-nivåer.
Kvalitetskontroll
Sterilt filtrerad
Förvaring och hållbarhetstid
Förvaras vid +2°C till +8°C, skyddad från ljus.
Efter öppnandet, förvara vid 4°C och använd inom 6-8 veckor.
Förhållanden vid leverans
Omgivande temperatur
Underhåll
Förvaras i kylskåp vid +2°C till +8°C i mörker. Undvik frysning och frekvent uppvärmning till +37°C, eftersom det försämrar produktkvaliteten.
Värm inte upp mediet över 37°C och använd inte okontrollerade värmekällor som t.ex. mikrovågsugnar.
Om endast en del av mediet ska användas, ta ut den mängd som behövs och värm det till rumstemperatur före användning.
Sammansättning
Kategori
Komponenter
Koncentration (mg/L)
Aminosyror
Glycin
18.75
L-Alanin
4.45
L-arginin HCl
147.50
L-Asparagin H₂O
7.50
L-Asparaginsyra
6.65
L-cystein HCl H₂O
17.56
L-cystin 2 HCl
31.29
L-glutaminsyra
7.35
L-glutamin
365.00
L-Histidin HCl H₂O
31.48
L-Isoleucin
54.47
L-Leucin
59.05
L-Lysin HCl
91.25
L-metionin
17.24
L-fenylalanin
35.48
L-prolin
17.25
L-Serin
26.25
L-Threonin
53.45
L-tryptofan
9.02
L-Tyrosin Dinatriumsalt
48.10
L-Valin
52.85
Vitaminer
D-Biotin
0.0035
Kolinklorid
8.98
D-kalciumpantotenat
2.24
Folsyra
2.66
myo-Inositol
12.60
Nikotinamid
2.02
Pyridoxin HCl
0.031
Pyridoxal HCl
2.00
Riboflavin
0.219
Tiamin HCl
2.17
Vitamin B12
0.68
Oorganiska salter
CaCl₂ 2 H₂O
154.50
CuSO₄ 5 H₂O
0.0013
Fe(NO₃)₃ 9 H₂O
0.05
FeSO₄ 7 H₂O
0.417
KCl
311.80
MgCl₂ 6 H₂O
61.20
MgSO₄
48.84
NaCl
6996.00
NaHCO₃
1200.00
Na₂HPO₄
71.02
NaH₂PO₄
54.30
ZnSO₄ 7 H₂O
0.432
Övriga komponenter
D-Glukos
3151.00
Hypoxantin
2.40
HEPES
3574.50
Linolsyra
0.042
Liponsyra
0.105
Fenolrött natriumsalt
8.63
Putrescin 2 HCl
0.081
Natriumpyruvat
55.00
Tymidin
0.365
, w: 3,0 g/L Glukos, w: stabil Glutamin, w: 2,0 mM Natriumpyruvat, w: 2,2 g/L NaHCO3")
- 0,02 vid 20-25 °C. Varje parti har testats för sterilitet och frånvaro av mykoplasma och bakterier. Förvaring Förvaras i kylskåp vid +2 °C till +8 °C i mörker. Frysning och uppvärmning till +37 °C försämrar produktens kvalitet. Värm inte mediet till mer än 37 °C och använd inte okontrollerbara värmekällor (t.ex. mikrovågsugnar). Om endast en del av mediet ska användas, ta ut denna mängd från flaskan och värm den till rumstemperatur. Hållbarhetstiden för alla medier utom basmediet är 6 till 8 veckor från öppningsdatumet. Sammansättning Komponenter mg/L Oorganiska salterKalciumklorid x 2H2O132,00 Magnesiumsulfat97,67 Kaliumklorid400 Natriumklorid6 460,00 Dinatriumvätefosfat (vattenfritt)504,00 Övriga komponenterD(+)-glukos (vattenfri)3 000 Glutation (reducerat)0 Köttpeptone600,00 Fenolrött natriumsalt11 AminosyrorL-alanin13 L-arginin x HCl42,1 L-asparagin x H2O45,03 L-asparaginsyra19,97 L-Cystein x HCl x H2O31,75 L-glutamin (stabil)219,15 L-glutaminsyra22,07 Glycin7,51 L-histidin x HCl x H2O20,96 L-hydroxiprolin19,67 L-isoleucin39,3 L-leucin39,36 L-lysin x HCl36,54 L-metionin14,92 L-fenylalanin16,52 L-prolin17,27 L-serin26,28 L-treonin17,87 L-tryptofan3,06 L-tyrosin dinatriumsalt26,10 L-valin17,57 Vitaminerp-Aminobensoesyra1,0 Askorbinsyra0,5 D(+)-biotin0,2 D-kalciumpantotenat0,2 Kolin klorid5 Folsyra10 Myo-inositol36 Nikotinamid0,5 Nikotinsyra0,5 Pyridoxal HCl0,5 Pyridoxin HCl0,50 Riboflavin0,2 Tiamin HCl0,2 Vitamin B122,0

Medium 199 erbjuder en rad olika tillämpningar inom området. Det kan effektivt upprätthålla kumulus-oocytkomplexet (COC) och stödja in vitro-mognaden av oocyter. Dessutom används det för att skölja aspirationskanaler vid insamling av ägg från tyska Holstein-kor. Medium 199 är dessutom ett utmärkt medium för odling av hjärtats endotelceller från råttor. Dessa tillämpningar visar på Medium 199:s mångsidighet och anpassningsförmåga till olika experimentella behov.
Mediets historia
Utvecklingen av Medium 199 på 1950-talet innebar ett betydande framsteg inom vävnadsodlingsmedier. Innan det introducerades var många odlingsmedier beroende av produkter och vävnadsextrakt från djur. Morgan och hans kollegor revolutionerade dock området genom att formulera en helt definierad näringskälla för cellkulturer. Genom sina experiment med olika kombinationer av vitaminer, aminosyror och andra faktorer upptäckte de de exceptionella tillväxtfrämjande egenskaperna hos Medium 199.
Kvalitetskontroll
pH = 7,2 +/
- 0,02 vid 20-25°C.
Varje parti har testats för sterilitet och frånvaro av mykoplasma och bakterier.
Förvaring
Förvaras i kylskåp vid +2°C till +8°C i mörker. Frysning och uppvärmning upp till +37°C minimerar produktens kvalitet.
Värm inte upp mediet till mer än 37° C och använd inte okontrollerbara värmekällor (t.ex. mikrovågsugnar).
Om endast en del av mediet ska användas, ta ut denna mängd ur flaskan och värm upp den i rumstemperatur.
Hållbarheten för alla medier utom basmediet är 8 veckor från tillverkningsdatumet.
Sammansättning
Komponenter
mg/L
Oorganiska salter
Kalciumklorid x 2H2O
264,92
Järn(III)nitrat x 9H2O
0,72
Magnesiumsulfat
97,67
Kaliumklorid
400,00
Natriumacetat x 3H2O
82,95
Natriumklorid
6,800.00
Natriumdivätefosfat x H2O
140,00
Övriga komponenter
Adeninsulfat
10,00
AMP
0,20
ATP
1,00
Kolesterol
0,20
2'-Deoxiribos
0,50
D(+)-Glukos vattenfri
1,000.00
Glutation (röd)
0,05
Guanin x HCl
0,30
Hypoxanthin
0,30
Fenol röd
10,00
D-Ribos
0,50
Tymin
0,30
Tween 80
4,90
Uracil
0,30
Xanthin
0,30
NaHCO3
2,200.00
Aminosyror
L-Alanin
25,00
L-arginin x HCl
70,00
L-Asparaginsyra
30,00
L-Cystein x HCl x H2O
0,10
L-Cystin
20,00
L-Glutamin stabil
149,00
L-glutaminsyra
67,00
Glycin
50,00
L-Histidin x HCl x H2O
21,88
L-Hydroxyprolin
10,00
L-Isoleucin
20,00
L-Leucin
60,00
L-Lysin x HCl
70,00
L-metionin
15,00
L-fenylalanin
25,00
L-prolin
40,00
L-Serin
25,00
L-Threonin
30,00
L-tryptofan
10,00
L-Tyrosin
40,00
L-Valin
25,00
Vitaminer
4-Amino bensoesyra
0,05
Askorbinsyra
0,05
D(+)-Biotin
0,01
Calciferol
0,10
D-kalciumpantotenat
0,01
Kolinklorid
0,50
Folsyra
0,01
myo-Inositol
0,05
Menadion
0,01
Nikotinsyra
0.025
Nikotinamid
0.025
Pyridoxal x HCl
0.025
Pyridoxol x HCl
0.025
Riboflavin
0,01
DL-α-tokoferolfosfat dinatriumsalt
0,01
Tiamin x HCl
0,01
Vitamin A-acetat
0,14

IMDM lämpar sig väl för snabbt växande cellkulturer med hög densitet, inklusive Jurkat-, COS-7
- och makrofagceller. De olika modifieringarna av IMDM som finns tillgängliga för en rad olika cellodlingsapplikationer kan hittas med hjälp av verktyget för medieväljare. Flytande medier tillhandahåller viktiga näringsämnen för alla cellodlingsapplikationer. Var och en av våra högkvalitativa cellodlingsmedier tillverkas enligt den ursprungligen publicerade formeln eller de modifieringar som krävs för att säkerställa konsekvent prestanda och stabilitet hos odlingsmediet.
IMDM jämfört med DMEM
IMDM innehåller kaliumnitrat i stället för järnnitrat samt HEPES och natriumpyruvat. De ytterligare komponenterna i IMDM gör det mer lämpligt för specialiserade celltyper och specifika tillämpningar än DMEM.
IMDM jämfört med RPMI
IMDM och RPMI har olika formuleringar som kan vara relevanta för stimulering med PMA/ionomycin. En betydande skillnad är koncentrationen av Ca2+. Medan RPMI innehåller 0,42 mM Ca2+, innehåller IMDM 1,49 mM.
Kvalitetskontroll
pH = 7,2 +/
- 0,02 vid 20-25°C.
Varje parti har testats för sterilitet och frånvaro av mykoplasma och bakterier.
Underhåll
Förvaras i kylskåp vid +2°C till +8°C i mörker. Frysning och uppvärmning upp till +37°C minimerar produktens kvalitet.
Värm inte upp mediet till mer än 37° C och använd inte okontrollerbara värmekällor (t.ex. mikrovågsugnar).
Om endast en del av mediet ska användas, ta ut denna mängd ur flaskan och värm upp den i rumstemperatur.
Hållbarheten för alla medier utom basmediet är 8 veckor från tillverkningsdatumet.
Sammansättning
Komponenter
mg/L
Oorganiska salter
Kalciumklorid x 2 H2O
219,00
Kaliumklorid
330,00
Kaliumnitrat
0.076
Vattenfritt magnesiumsulfat
97,73
Natriumklorid
4,505.00
Natriumdivätefosfat vattenfritt
109,00
Natriumselenit
0,02
Övriga komponenter
D(+)-Glukos vattenfri
4,500.00
HEPES
5,958.00
Natriumpyruvat
110,00
Fenolrött
15,00
Aminosyror
L-Alanin
25,00
L-arginin x HCl
84,00
L-Asparagin x H2O
25,00
L-Asparaginsyra
30,00
L-kystin x 2HCl
91,24
L-Glutamin
584,00
L-glutaminsyra
75,00
Glycin
30,00
L-Histidin x HCl x H2O
42,00
L-Isoleucin
104,80
L-Leucin
104,80
L-Lysin x HCl
146,20
L-metionin
30,00
L-fenylalanin
66,00
L-prolin
40,00
L-Serin
42,00
L-Threonin
95,20
L-tryptofan
16,00
L-Tyrosin x 2Na
104,20
L-Valin
93,60
Vitaminer
D(+)-Biotin
0.013
D-kalciumpantotenat
4,00
Kolinklorid
4,00
Folsyra
4,00
myo-Inositol
7,20
Medier för cellodling: En översikt
Inom biovetenskaperna är cellodling en av de viktigaste metoderna. Uttrycket "cellodling" betyder att celler, vävnader eller organ tas ut från ett djur eller en växt och sedan implanteras i en artificiell miljö som är gynnsam för deras överlevnad och/eller tillväxt De grundläggande miljöbehoven för optimal cellutveckling är kontrollerad temperatur, ett substrat för cellfästning, ett adekvat tillväxtmedium och en inkubator som upprätthåller optimalt pH och osmolalitet. Cellerna måste ha dessa förutsättningar för att kunna växa till sin fulla potential.
Valet av ett lämpligt tillväxtmedium för in vitro-odling är det steg i cellodlingen som är både det mest kritiska och det mest avgörande. Ett tillväxtmedium, även kallat odlingsmedium, är en vätska eller gel som är formulerad för att uppmuntra organismers utveckling i mikroskopisk, cellulär eller växtliknande skala. Det medium som används för att odla celler innehåller ofta en tillräcklig mängd energi och ämnen som styr cellcykeln. De viktigaste komponenterna i ett odlingsmedium är aminosyror, vitaminer, oorganiska salter, glukos och serum. Serumet tillsätts till mediet eftersom det fungerar som en källa till tillväxtfaktorer, hormoner och bindningsfaktorer. Förutom att tillhandahålla näringsämnen bidrar mediet också till att upprätthålla pH- och osmolalitetsnivåerna.
Olika typer av medium som används vid cellodling
Både mänskliga och animaliska celler kan odlas i antingen ett artificiellt eller syntetiskt medium eller i ett helt naturligt medium som kompletteras med naturliga element. Nedan följer en översikt över de olika typer av medier som finns tillgängliga för närvarande.
Naturliga medier
Endast biologiska vätskor som existerar i sitt naturliga tillstånd kan hittas i naturliga medier. Naturliga medier är mycket användbara och enkla för odling av en mängd olika typer av djurceller. Bristen på förståelse för de exakta komponenter som ingår i naturliga medier är den främsta faktorn som bidrar till den låga repeterbarheten hos de resultat som erhålls med hjälp av naturliga medier.
Artificiella medier
Vid framställning av artificiella eller syntetiska medier tillsätts näringsämnen (både organiska och oorganiska), serumproteiner, kolhydrater, kofaktorer, vitaminer och salter samt gasfaserna O2 och CO2 [1].
Olika typer av artificiella medier har utvecklats för att uppfylla en eller flera av följande funktioner 1) Omedelbar överlevnad (en balanserad saltlösning med exakt pH-värde och osmotiskt tryck). 2) Förlängd överlevnad (en balanserad saltlösning kompletterad med olika formuleringar av organiska kemikalier och/eller serum). 3) Utveckling på obestämd tid. 4) Specialiserade funktioner.
Det finns fyra olika klassificeringar för artificiella medier:
Serumhaltiga medier
Den vanligaste typen av tillsats som finns i medium som används för odling av djurceller är fetalt bovint serum. Det tillsätts till odlingsmediet som ett billigt komplement för att uppnå bästa möjliga tillväxtförhållanden. Förutom att serumet fungerar som transportör eller kelator för näringsämnen som är instabila eller vattenolösliga, hormoner och tillväxtfaktorer, proteashämmare och andra ämnen, binder och neutraliserar det också skadliga molekyler.
Serumfritt medium
Förekomsten av serum i medierna har ett antal nackdelar och kan orsaka stora tolkningsfel inom immunologisk forskning [2, 3]. Det har skapats en mängd olika serumfria medier [4, 5]. Dessa medier är i allmänhet specifikt formulerade för att stödja odlingen av en enda celltyp, såsom Knockout Serum Replacement och Knockout DMEM från Thermo Fisher Scientific, och mTESR medium från Stem Cell Technologies [6], för stamceller [7].
Dessutom innehåller dessa medier definierade mängder av renade tillväxtfaktorer, lipoproteiner och andra proteiner, som annars vanligtvis tillförs via serum [8]. Dessa medier kallas ofta "definierade kulturella medier" eftersom de komponenter som ingår i dessa medier är väl kända.
Kemiskt definierade medier
Dessa medier innehåller ultrarena oorganiska och organiska komponenter som inte har förorenats av någon form av kontaminering. De kan också innehålla rena proteintillsatser, t.ex. tillväxtfaktorer.
den genetiska modifieringen av bakterier eller jäst, tillsammans med tillsats av särskilda fettsyror, vitaminer, kolesterol och aminosyror, resulterar i produktion av deras beståndsdelar [9].
Proteinfria medier
Proteinfria medier är sådana som inte innehåller något protein alls utan endast icke-proteinbaserade element. Jämfört med medier med tillsatt serum främjar användningen av medier utan tillsatt protein större cellproliferation och proteinuttryck och gör det lättare att rena eventuella produkter som genereras i en nedströmsprocess [10-12]. Protein ingår inte i formuleringar som MEM och RPMI-1640. Ett proteintillskott kan dock administreras om det är nödvändigt.
Odlingsmedium och dess grundläggande komponenter
Kommersiella odlingsmedier kan köpas som pulver eller vätska och innehåller ofta en mängd olika näringsämnen som aminosyror, glukos, salter, vitaminer och andra kosttillskott.
Behoven av dessa komponenter är olika för varje cellinje, och dessa variationer är orsaken till det stora antalet olika formuleringar av medier. Varje komponent ansvarar för en viss funktion, som kommer att beskrivas i följande stycken:
System för buffring
För att upprätthålla optimala odlingsförhållanden måste pH-värdet kontrolleras, vilket ofta görs med hjälp av ett av två buffringssystem:
Naturligt buffringssystem
CO2/H2CO3-förhållandet i atmosfären är lika stort som i mediet, vilket skapar en naturlig buffertmekanism. För att bevara den naturliga buffertmekanismen måste kulturer förvaras i en luftmiljö med 5-10% CO2, vilket ofta uppnås genom att använda en CO2-inkubator. En av de bästa sakerna med att använda en naturlig buffert är hur billig och säker den är.
HEPES
Kemisk buffring med hjälp av zwitterjonen HEPES har en större buffringsförmåga i pH-området 7,2-7,4 och behöver inte en reglerad gasformig miljö. För vissa celltyper kan en större dos av HEPES vara skadlig. Medier som innehåller HEPES är också mycket mer känsliga för de fototoxiska effekterna av fluorescerande ljus [13].
Fenolrött
PH-indikatorn fenolrött ingår ofta i kommersiellt tillgängliga odlingsmedier, vilket möjliggör kontinuerlig övervakning av pH. Genom att cellerna expanderar orsakar de metaboliter som produceras av dessa celler en förändring av pH-värdet och därmed en färgförändring i mediet. Fenolrött har en dubbel effekt på ett mediums färg, det blir gult vid surt pH och lila vid alkaliskt pH. pH 7,4, det optimala värdet för cellodling, gör att mediet blir fluorescerande rött.
Men fenolrött har några nackdelar: För det första kan fenolrött simulera effekten av ett antal steroidhormoner, främst östrogen [14]. Vid studier av östrogenkänsliga celler, t.ex. bröstvävnad, rekommenderas därför ett medium fritt från fenolrött. Natrium-kaliumbalansen störs av förekomsten av fenolrött i flera serumfria formuleringar. Tillsats av serum eller bovint hypofyshormon till mediet kan motverka denna effekt [15]. För det tredje försvåras detektion i flödescytometriska experiment av förekomsten av fenolrött.
Oorganiska salter
Medier som innehåller oorganiska salter, t.ex. natrium-, kalium- och kalciumjoner, bidrar till att upprätthålla osmotisk jämvikt och reglera membranpotentialen.
Aminosyror
Eftersom aminosyror är de grundläggande komponenterna i protein är de en viktig komponent i varje cellodlingsmedium som någonsin har skapats. Eftersom cellerna inte kan producera vissa aminosyror på egen hand är det viktigt att odlingsmediet innehåller essentiella aminosyror. De är nödvändiga för cellernas spridning och koncentrationen av dem avgör vilken maximal celldensitet som kan uppnås. I synnerhet är L-glutamin, en essentiell aminosyra, särskilt viktig.
L-glutamin fungerar som en sekundär energikälla för ämnesomsättningen och bidrar med kväve till produktionen av NAD, NADPH och nukleotider. Eftersom L-glutamin är en instabil aminosyra som med tiden övergår i en form som cellerna inte kan tillgodogöra sig, måste den tillföras mediet.
Dessutom kan icke-essentiella aminosyror tillföras mediet för att fylla på de aminosyror som har förbrukats under tillväxtprocessen. Tillväxten av cellerna stimuleras och deras livskraft ökar när tillväxtmediet kompletteras med icke-essentiella aminosyror.
Kolhydrater
Kolhydrater i form av sockerarter är den viktigaste energikällan. Många av medierna innehåller också maltos och fruktos utöver de vanligare sockerarterna glukos och galaktos.
Proteiner och peptider
Albumin, transferrin och fibronektin är de vanligaste proteinerna och peptiderna. De är särskilt viktiga i medier som inte innehåller serum. Albumin, transferrin, aprotinin, fetuin och fibronektin är några av de proteiner som kan finnas i serum, som är en rik källa till protein.
Albumin är det primära proteinet i blodet och dess funktion är att binda och transportera olika ämnen, bland annat vatten, salter, fria fettsyror, hormoner och vitaminer, mellan olika organ och celler. Albuminets förmåga att binda till sig kemikalier gör det till en effektiv kandidat för att avlägsna skadliga föreningar från det medium som celler odlas i.
Aprotinin är ett skyddande medel i cellodlingssystem, eftersom det är stabilt vid neutralt och surt pH samt motståndskraftigt mot höga temperaturer och den förstörelse som kan orsakas av proteolytiska enzymer. Det kan hämma ett antal serinproteaser, bland annat trypsin.
Fetuin är ett glykoprotein som kan påvisas i större mängder i serum från foster och nyfödda djur jämfört med serum från vuxna. Dessutom fungerar det som en serinproteashämmare. Proteinet fibronektin är en viktig komponent i processen för celladhesion. Transferrin är ett protein som transporterar järn och ansvarar för att leverera järn till cellernas membran.
Fettsyror och lipider
De spelar en avgörande roll i serumfritt medium när serum saknas.
Vitaminer
Många vitaminer är nödvändiga för cellutveckling och cellproliferation. Vitaminer kan inte produceras i tillräcklig mängd av cellerna och är därför nödvändiga som kosttillskott i vävnadskulturer.
I cellkultur är serum den primära källan till vitaminer, men medier behandlas också med olika vitaminer för att göra dem lämpliga för en viss celltyp. Vanligtvis används B-vitaminer för att stimulera tillväxt.
Spårämnen
Kemiska grundämnen som koppar, zink, selen och tricarboxylsyraintermediärer kallas spårämnen. Spårämnen tillsätts ofta i medier som inte innehåller serum för att ersätta de ämnen som normalt finns i serum. Dessa element är viktiga kemiska komponenter som krävs för en sund cellutveckling. Många biokemiska reaktioner är beroende av vissa mikronäringsämnen, t.ex. enzymaktivitet.
Tillskott av medium
Det fulla tillväxtmedium som föreslås för vissa cellinjer behöver extra komponenter som saknas i baslinjemediet och serumet. Dessa kosttillskott stöder celltillväxt och lämplig metabolisk funktion.
Även om hormoner, tillväxtfaktorer och signalmolekyler är nödvändiga för en lämplig spridning av vissa cellinjer, bör följande försiktighetsåtgärder alltid vidtas: Eftersom tillsats av kosttillskott kan förändra osmolaliteten i det kompletta tillväxtmediet, vilket kan hämma cellutvecklingen, är det alltid lämpligt att kontrollera osmolaliteten efter tillsats av kosttillskott. För de flesta cellinjer ligger den optimala osmolaliteten mellan 260 och 320 mOSM/kg.
Antibiotika
Antibiotika används ofta för att hämma utvecklingen av bakterie- och svampföroreningar [16], även om de inte är nödvändiga för celltillväxt. Eftersom antibiotika kan dölja kontaminering med mykoplasma och resistenta bakterier rekommenderas inte rutinmässig användning av antibiotika för cellodling [17, 18].
Dessutom kan antibiotika störa metabolismen i överkänsliga celler. Penicillin-streptomycin-kombinationerna från MilliporeSigma och Life Technologies används ofta. Plasmocin har använts vid odling av gliomcellinjerna TS603, TS516 och BT260 [19], och det har visat sig vara effektivt för att avlägsna mykoplasmakontaminering (20).
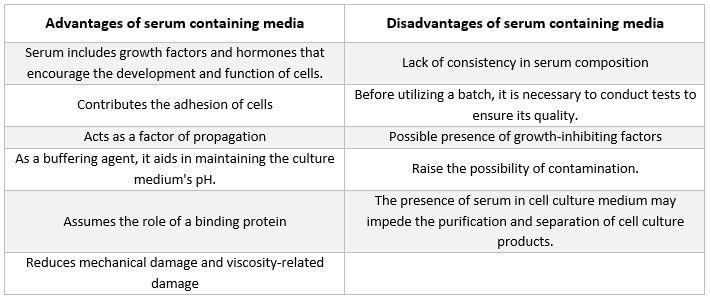
Serum
Albuminer, tillväxtfaktorer och tillväxthämmare finns alla i serum. Serum är en av de viktigaste komponenterna i cellodlingsmediet eftersom det innehåller aminosyror, proteiner, vitaminer (särskilt fettlösliga vitaminer som A, D, E och K), kolhydrater, lipider, hormoner, tillväxtfaktorer, mineraler och spårämnen.
Serum från foster och kalv från nötkreatur används ofta för att främja utvecklingen av odlade celler. Fetalt serum innehåller rikligt med tillväxtfaktorer och lämpar sig för cellkloning och utveckling av känsliga celler. På grund av dess minskade tillväxtfrämjande förmåga används kalvserum i kontaktinhiberingsexperiment. Normala tillväxtmedier innehåller ofta 2-10% serum. Tillsats av serum till odlingsmedium har följande syften [21]:
-
Serumet levererar viktiga näringsämnen till cellerna (både i lösning och bundna till proteiner).
-
Flera tillväxtfaktorer och hormoner som är involverade i tillväxtfrämjande och specialiserad cellaktivitet ingår i serum.
-
Det innehåller många bindande proteiner, som albumin och transferrin, som transporterar andra kemikalier in i cellen. Albumin transporterar t.ex. fetter, vitaminer och hormoner in i cellerna.
-
Det innehåller också proteiner, t.ex. fibronektin, som ökar cellernas vidhäftning till underlaget. Dessutom producerar det spridningselement som hjälper till vid cellutvidgning före delning.
-
Det levererar proteashämmare som förhindrar proteolys i celler.
-
Den innehåller också mineraler som Na+, K+, Zn2+ och Fe2+.
-
Det ökar viskositeten i mediet, vilket skyddar cellerna från mekanisk skada under omrörning av suspensionskulturen.
-
Det är också en buffert.
Referenser
[1] Morgan J, Morton H, Parker R. Nutrition av djurceller i vävnadsodling; inledande studier av ett syntetiskt medium. Proc Soc Exp Biol Med. 1950;73:1-8
[2] Kerbel R, Blakeslee D. Snabb adsorption av en komponent i serum från kalvfoster av däggdjursceller i kultur. En potentiell källa till artefakter i studier av antisera mot cellspecifika antigener. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. Tillsats av serum till det medium som används för beredning av cellsuspensioner som en möjlig källa till artefakter i cellmedierade reaktioner som studeras med hjälp av popliteal lymfkörteltest. J Immunogenet. 1980;7:483-9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Kommersiella serumfria medier: hybridomtillväxt och produktion av monoklonala antikroppar. J Immunol Methods. 1991;145:175-83
[5] Barnes D, Sato G. Metoder för tillväxt av odlade celler i serumfritt medium. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O,et al. HSP70 chaperonerar RNA-fri TDP-43 till anisotropiska intranukleära flytande sfäriska skal. Vetenskap. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J,et al. Down-syndrome-inducerad senescens stör den nukleära arkitekturen hos neurala progenitorer. Cell Stem Cell. 2022;29:116-130.e7
[8] Iscove N, Melchers F. Fullständig ersättning av serum med albumin, transferrin och sojabönlipid i kulturer av lipopolysackaridreaktiva B-lymfocyter. J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Systematisk förbättring av ett kemiskt definierat proteinfritt medium för hybridomtillväxt och produktion av monoklonala antikroppar. J Biotechnol. 1996;45:111-23
[10] Darfler F. Ett proteinfritt medium för tillväxt av hybridomer och andra celler i immunsystemet. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Serumfri cellodling: en enhetlig metod. Cell. 1980;22:649-55
[12] Hamilton W, Ham R. Klonal tillväxt av cellinjer från kinesisk hamster i proteinfria medier. In Vitro. 1977;13:537-47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Analys av de cytotoxiska effekterna av ljusexponerat HEPES-innehållande odlingsmedium. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. Fenolrött i vävnadskulturmedier är ett svagt östrogen: konsekvenser för studien av östrogenresponsiva celler i kultur. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Utveckling av serumfria medier. I: Master JRW, redaktör. Animal Cell culture, 3:e upplagan. Oxford: Oxford University Press; 2000.
[16] Perlman D. Användning av antibiotika i cellodlingsmedier. Metoder Enzymol. 1979;58:110-6
[17] McGarrity G. Spridning och kontroll av mykoplasmainfektion i cellkulturer. In Vitro. 1976;12:643-8
[18] Masters J, Stacey G. Byte av medium och passering av cellinjer. Nat protokoll. 2007;2:2276-84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M,et al. Histondemetylas KDM6A känner direkt av syre för att kontrollera kromatin och cellöde. Vetenskap. 2019;363:1217-1222
[20] Molla Kazemiha V, Azari S, Amanzadeh A, Bonakdar S, Shojaei Moghadam M, Habibi Anbouhi M,et al. Efficiency of Plasmocin™ on various mammalian cell lines infected by mollicutes in comparison with commonly used antibiotics in cell culture: a local experience. Cytoteknologi. 2011;63:609-20
[21] Kragh Hansen U. Molekylära aspekter av ligandbindning till serumalbumin. Pharmacol Rev. 1981;33:17-53