
, L-alanyl-L-glutamine-oplossing")
Volume: 100 ml
Opslag: ≤ –15 °C
Steriliteit: Steriel gefilterd
Stabiele glutamineoplossing (L-Alanyl-L-Glutamine, 200 mM) is een zeer stabiele dipeptidevorm van L-glutamine, ontworpen als directe vervanging voor conventionele L-glutamine in celkweekmedia. L-glutamine is een essentieel aminozuur en een belangrijke energiebron voor gekweekte cellen, en speelt een cruciale rol in celgroei, metabolisme en eiwitsynthese.
Toepassing en voordelen
In standaard vloeibare media wordt L-glutamine bij 37 °C relatief snel afgebroken, wat leidt tot de vorming van toxische bijproducten zoals ammoniumionen die de levensvatbaarheid van cellen en de experimentele resultaten negatief kunnen beïnvloeden. Stabiele glutamine overwint deze beperking door een niet-afbreekbare dipeptidevorm te bieden die intact blijft in kweekomstandigheden.
Cellen splitsen de dipeptidebinding enzymatisch om L-glutamine vrij te maken wanneer dat nodig is, waardoor een continue verse toevoer wordt gegarandeerd en de ophoping van schadelijke afvalproducten wordt voorkomen. Dit maakt de oplossing bijzonder voordelig voor langdurige celculturen en groeisystemen met een hoge dichtheid.
Samenstelling en gebruik
De L-Alanyl-L-Glutamine-oplossing wordt bereid in water van celkweekkwaliteit met een concentratie van 200 mM en wordt steriel gefilterd voor toepassingen die gevoelig zijn voor besmetting. Het kan direct worden verdund in complete media volgens de experimentele vereisten. Bewaar bij ≤ –15 °C en vermijd herhaalde vries-dooicycli om de productstabiliteit te behouden.
Uitsluitend voor onderzoeksdoeleinden. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
")
Inhoud: 100 ml
Opslag: +2 °C tot +8 °C
Steriliteit: Steriel gefilterd
HEPES-bufferoplossing (1 M), ook bekend als N-2-hydroxyethylpiperazine-N-2-ethaansulfonzuur, is een zwitterionisch organisch bufferingsmiddel dat veel wordt gebruikt in celkweekmedia. Het is ontworpen om stabiele pH-waarden te handhaven binnen het fysiologische bereik van 6,7 tot 8,6, waardoor een optimale celfunctie tijdens in-vitro-toepassingen wordt ondersteund.
Toepassing en voordelen
HEPES biedt betrouwbare buffercapaciteit in celkweeksystemen, met name wanneer cellen buiten een CO₂-incubator worden behandeld. De toevoeging van 10-25 mM HEPES aan kweekmedia zorgt voor een verbeterde pH-stabiliteit tijdens langdurige manipulatieperioden, waardoor consistente experimentele omstandigheden kunnen worden gehandhaafd.
Deze buffer is membraanondoorlatend, heeft minimale invloed op biochemische reacties en vertoont een sterke chemische en enzymatische stabiliteit. Deze eigenschappen maken het geschikt voor een breed scala aan celkweek
- en biochemische toepassingen.
Samenstelling en gebruik
De oplossing wordt geleverd als een 1 M concentraat, bereid in water van celkweekkwaliteit, en is steriel gefilterd voor gebruik in omgevingen die gevoelig zijn voor besmetting. Het kan worden verdund tot de gewenste werkconcentratie, afhankelijk van de toepassingsvereisten. Bewaren bij +2 °C tot +8 °C en hanteren onder aseptische omstandigheden om de integriteit van het product te behouden.
Uitsluitend voor onderzoeksdoeleinden. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
, 250 g/l")
Inhoud: 50 ml
Bewaring: +2 °C tot +8 °C
Steriliteit: Steriel gefilterd
D-(+)-glucose-oplossing (dextrose-oplossing) is een steriel, gebruiksklaar supplement dat de natuurlijk voorkomende suiker D-(+)-glucose bevat, een centraal onderdeel van het celmetabolisme. Glucose is betrokken bij essentiële biologische processen zoals energieproductie, glycosylatie en de vorming van glycanen die bijdragen aan de celstructuur en -functie.
Toepassing en voordelen
Deze glucoseoplossing wordt veel gebruikt als supplement in celkweekmedia en in tal van cellulaire en moleculaire biologische toepassingen. Als primaire koolstof
- en energiebron ondersteunt glucose de celgroei, proliferatie en metabolische activiteit. Door zijn betrokkenheid bij biosynthetische routes is het ook van cruciaal belang voor het behoud van een normale cellulaire fysiologie en experimentele consistentie.
Samenstelling en gebruik
De oplossing wordt geleverd in een hoge concentratie van 250 g/l glucose, waardoor deze flexibel kan worden verdund in kweekmedia volgens de experimentele vereisten. De oplossing is steriel gefilterd om geschikt te zijn voor toepassingen waarbij besmetting een risico vormt. Bewaar bij +2 °C tot +8 °C en behandel aseptisch om de productkwaliteit en stabiliteit te behouden.
Uitsluitend voor onderzoeksdoeleinden. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
, oplossing 100x")
Inhoud: 10 ml
Opslag: +2 °C tot +8 °C
Steriliteit: Steriel gefilterd
Insuline-Transferrine-Selenium (ITS) Oplossing (100x) is een chemisch gedefinieerd supplement dat is ontworpen voor een breed scala aan celkweektoepassingen. Het wordt meestal gebruikt als additief voor basale celkweekmedia om de celgroei te ondersteunen onder omstandigheden met verminderd serum of zonder serum.
Toepassing en voordelen
Ons ITS-supplement levert essentiële componenten die nodig zijn voor de optimale werking van serumvrije media. Door conventionele voedingsmedia aan te vullen met ITS kan de behoefte aan foetaal runderserum (FBS) voor het routinematig onderhoud van veel cellijnen aanzienlijk worden verminderd. Dit helpt de variabiliteit die gepaard gaat met het gebruik van serum te minimaliseren, terwijl een consistente celgroei en levensvatbaarheid behouden blijven.
Insuline ondersteunt de opname en het metabolisme van belangrijke voedingsstoffen door de cellen, transferrine bevordert het ijzertransport en selenium draagt bij aan de antioxidantafweer en enzymatische activiteit. Samen bevorderen deze componenten een evenwichtig celmetabolisme en een verbeterde reproduceerbaarheid in gedefinieerde kweeksystemen.
Samenstelling en gebruik
Insuline-Transferrine-Selenium (ITS) wordt geleverd als een 100x geconcentreerde oplossing in Earle's Balanced Salt Solution (EBSS) zonder fenolrood. Voor standaardtoepassingen verdunt u het 1:100 in het geschikte basismedium om de aanbevolen werkconcentratie te bereiken. Bewaar bij +2 °C tot +8 °C en hanteer onder aseptische omstandigheden om de productstabiliteit en steriliteit te behouden.
Uitsluitend voor onderzoeksdoeleinden. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
")
Volume: 5 ml
Opslag: +2 °C tot +8 °C
Steriliteit: Steriel gefilterd
Recombinant humaan insulineoplossing is een chemisch gedefinieerd supplement dat vaak wordt gebruikt voor de kweek van zoogdiercellijnen, waaronder Chinese hamsterovariumcellen (CHO-cellen). Deze oplossing van celkweekkwaliteit bevat recombinant humaan insuline dat tot expressie is gebracht in Saccharomyces cerevisiae, wat een hoge zuiverheid en consistente prestaties bij onderzoekstoepassingen garandeert.
Toepassing en voordelen
Insuline wordt routinematig toegevoegd aan serumvrije en chemisch gedefinieerde media om de celgroei en productiviteit te bevorderen. Als belangrijk regulerend hormoon ondersteunt insuline de opname, het gebruik en de opslag van glucose, aminozuren en vetzuren door de cellen. Het remt ook de afbraak van glycogeen, eiwitten en lipiden, wat bijdraagt aan een verbeterde levensvatbaarheid van de cellen en metabolische stabiliteit in kweeksystemen. De chemisch gedefinieerde formulering ondersteunt de reproduceerbaarheid en minimaliseert variabiliteit in gevoelige celkweekprocessen.
Biologische eigenschappen en gebruik
Insuline is een polypeptidehormoon met twee ketens dat van nature wordt geproduceerd door de β-cellen van de pancreaseilandjes. Het heeft een molecuulgewicht van ongeveer 5.800 Da. De α
- en β-ketens zijn verbonden door twee disulfidebindingen tussen de ketens, en de α-keten bevat één disulfidebinding binnen de keten. Voor celkweektoepassingen moet de oplossing onder aseptische omstandigheden worden behandeld en bij +2 °C tot +8 °C worden bewaard om de stabiliteit en prestaties te behouden.
Uitsluitend voor onderzoeksdoeleinden. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
")
Inhoud: 100 ml
Bewaring: +2 °C tot +8 °C
Steriliteit: Steriel gefilterd
Natriumpyruvaatoplossing (100 mM) is een steriel, gebruiksklaar supplement dat is ontwikkeld om een extra, gemakkelijk toegankelijke energiebron voor celkweekmedia te bieden. Natriumpyruvaat speelt een belangrijke rol in het cellulaire energiemetabolisme en ondersteunt de groei van metabolisch actieve en snel prolifererende cellen, zoals tumorcellen. Suppletie kan de levensvatbaarheid van cellen verbeteren en helpen de metabolische stabiliteit in kweeksystemen te behouden.
Toepassing en voordelen
Deze oplossing wordt veel gebruikt in routinematige celculturen om media te verrijken met pyruvaat en optimale groeiomstandigheden te bevorderen. Het ondersteunt de ATP-productie, kan oxidatieve stress helpen verminderen en draagt bij aan een verbeterde metabolische prestatie van gekweekte cellen. Het product wordt vervaardigd in water van celkweekkwaliteit en is steriel gefilterd, waardoor een consistente kwaliteit en reproduceerbaarheid in onderzoeksworkflows wordt gegarandeerd.
Gebruik en compatibiliteit
De aanbevolen eindconcentratie voor de meeste celkweektoepassingen is 1 mM natriumpyruvaat, te verkrijgen door de 100 mM voorraadoplossing 1:100 te verdunnen in compleet kweekmedium. De oplossing is compatibel met een breed scala aan basismedia en zoogdiercellijnen. Bewaren bij +2 °C tot +8 °C en beschermen tegen besmetting om de productstabiliteit te behouden.
Alleen voor onderzoeksdoeleinden. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
")
Volume: 100 ml Opslag: ≤-15°C Steriliteit: Steriel gefilterd
Antibiotic/Antimycotic Solution (100x) is een steriel, gebruiksklaar concentraat dat is ontworpen om de risico's van microbiële besmetting in celkweken en aanverwante laboratoriumtoepassingen te verminderen. Deze 100x-oplossing bevat een beproefde combinatie van penicilline, streptomycine en amfotericine B, die een breed spectrum antimicrobiële activiteit biedt tegen Gram-positieve en Gram-negatieve bacteriën, gisten en filamenteuze schimmels. De formulering is geschikt voor gebruik in eukaryote celculturen, bacteriële media en andere besmettingsgevoelige systemen, ter ondersteuning van schone en consistente laboratoriumwerkzaamheden.
Toepassing en voordelen Deze oplossing is geoptimaliseerd voor routinematige onderzoeksprotocollen en wordt veel gebruikt om aseptische omstandigheden in celkweekworkflows te handhaven. Het biedt betrouwbare prestaties in contaminatiegevoelige omgevingen en helpt onderzoekers het risico van microbiële overgroei te verminderen zonder de gezondheid van de cellen of de experimentele reproduceerbaarheid in gevaar te brengen. De steriel gefilterde formulering elimineert de noodzaak voor extra solubilisatiestappen, ondersteunt een gestroomlijnde mediabereiding en vermindert de variabiliteit in dagelijkse labprocedures.
Gebruik en compatibiliteit Om standaard werkconcentraties te bereiken, verdunt u de oplossing 1:100 in uw volledige kweekmedium. Het product is compatibel met een breed scala aan zoogdiercellijnen en basismedia. Door de consistente beschikbaarheid van de voorraad profiteren onderzoekers van een betrouwbare leveringscontinuïteit en vereenvoudigde logistieke planning. De oplossing moet worden bewaard bij ≤ -15 °C en worden beschermd tegen herhaalde vries-dooicycli om de stabiliteit te behouden. Alleen voor onderzoek. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.
")
Volume: 100 ml Opslag: +2°C tot +8°C Steriliteit: Steriel gefilterd
MEM Non-Essential Amino Acids (100x) is een steriel supplement ontworpen om de celgroei en levensvatbaarheid in zoogdiercelkweeksystemen te verbeteren. De formulering komt overeen met een 100x concentraat van de niet-essentiële aminozuren die voorkomen in standaard Minimum Essential Medium (MEM), waardoor directe aanvulling van basismedia met minimale voorbereiding mogelijk is.
Toepassing en voordelen Dit supplement biedt een extra aminozuurpool voor snel prolifererende cellen of voor cellijnen die het vermogen hebben verloren om niet-essentiële aminozuren de novo te synthetiseren. Door de metabolische belasting van biosynthese te verlichten, ondersteunt het verbeterde groeikinetiek, verlengde levensvatbaarheid en grotere experimentele consistentie, vooral in culturen die gevoelig zijn voor voedingsstoffen of culturen met een hoge dichtheid.
Samenstelling en gebruik De oplossing bevat glycine, L-alanine, L-asparagine, L-asparaginezuur, L-glutaminezuur, L-proline en L-serine. Het is compatibel met MEM en de meeste andere standaardmedia. Verdun voor gebruik 1:100 in het uiteindelijke kweekmedium. Dit product is steriel gefilterd en klaar voor gebruik zonder extra stappen. Alleen voor onderzoek. Niet voor gebruik in diagnostische of therapeutische procedures. Niet voor gebruik bij mensen of dieren.

Accutase is een gebruiksklare, steriel gefilterde celafhechtingsoplossing die is ontworpen als een zacht alternatief voor trypsine/EDTA voor het losmaken van hechtende zoogdiercellen van standaard plastic weefselkweekmateriaal en met adhesie gecoate oppervlakken. Het combineert proteolytische en collagenolytische enzymactiviteit in een uitgebalanceerde zoutoplossing om een effectieve maar gecontroleerde dissociatie te bieden, waarbij eiwitten op het celoppervlak behouden blijven en een hoge levensvatbaarheid na passage en snelle herhechting worden ondersteund.
De Accutase-formule is gebaseerd op Dulbecco's fosfaatgebufferde zoutoplossing (DPBS) met EDTA en fenolrood als visuele pH-indicator. De enzymen zijn van niet-zoogdierlijke en niet-bacteriële oorsprong, waardoor Accutase bijzonder geschikt is voor stamcelonderzoek, vaccinworkflows en elke toepassing waarbij verontreinigingen van dierlijke of microbiële oorsprong tot een minimum moeten worden beperkt. De oplossing remt zichzelf bij 37 °C, zodat er na het losmaken geen neutraliserend reagens of serumhoudend medium nodig is – cellen kunnen direct in vers medium worden overgebracht.
Belangrijkste kenmerken
Kant-en-klare 1x steriel gefilterde vloeistof – verdunning of reconstitutie is niet nodig
Gecombineerde proteolytische en collagenolytische enzymactiviteit voor zachte dissociatie
Elke batch is gestandaardiseerd op een gedefinieerde dissociatieactiviteit voor consistentie tussen de verschillende partijen
Enzymen van niet-zoogdierlijke en niet-bacteriële oorsprong
Auto-inhibitie bij 37 °C – geen neutraliserende oplossing nodig
Geformuleerd in Dulbecco’s PBS met EDTA
Fenolrood toegevoegd als visuele pH-indicator
pH 6,8 – 7,8
Typische toepassingen
Accutase dissocieert op milde wijze een breed scala aan hechtende en gevoelige celtypen, waaronder menselijke embryonale stamcellen (hESCs), menselijke geïnduceerde pluripotente stamcellen (iPSCs), neurale stamcellen, primaire neuronen en routinematig gekweekte hechtende lijnen zoals HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 en A549. Typische toepassingen zijn onder meer:
Routinematige subcultuur en passage van hechtende zoogdiercellen
Zachte dissociatie van afzonderlijke cellen van hESC's, iPSC's en andere gevoelige lijnen
Monstervoorbereiding voor flowcytometrie en FACS-analyse
Analyse van celoppervlaktemarkers waarbij de integriteit van het epitoop van belang is
Testen op celmigratie, proliferatie en apoptose
Rusttoestandtesten door serumuitputting en oncogeentransfectiestudies
Migratietests voor tumorcellen en neurale crestcellen
Opschaling van de productie in bioreactor-workflows
Voor routinewerkzaamheden brengt u ongeveer 10 ml Accutase per 75 cm2 kweekoppervlak aan en incubeert u dit gedurende 5–10 minuten bij kamertemperatuur. De optimale incubatietijd moet voor elke cellijn worden bepaald en mag niet langer zijn dan een uur. Spoel de cellaag vóór toevoeging af met een Ca2+/Mg2+-vrije zoutoplossing, zoals DPBS zonder calcium en magnesium, om restserum en tweewaardige kationen te verwijderen.
Behandeling en opslag
Bewaar de ongeopende fles ingevroren bij -15 °C of lager. Ontdooi bij kamertemperatuur of 's nachts bij +2 °C tot +8 °C. Ontdooi Accutase niet in een waterbad van 37 °C, aangezien verhoogde temperaturen de enzymactiviteit verminderen. Na ontdooien kan de oplossing maximaal 2 maanden worden bewaard bij +2 °C tot +8 °C; bewaar niet bij kamertemperatuur. Verwarm het reagens niet voor op 37 °C vóór gebruik – voeg het direct toe aan gewassen cellen bij kamertemperatuur. Voor een lange houdbaarheid wordt het verdelen in porties voor eenmalig gebruik aanbevolen om herhaalde ontdooicycli te voorkomen. Werk altijd onder aseptische omstandigheden.
Kwaliteit
Vervaardigd volgens strenge kwaliteitsnormen. Elke batch Accutase wordt steriel gefilterd en getest op steriliteit, pH, uiterlijk en dissociatieactiviteit om consistente, reproduceerbare prestaties van batch tot batch te garanderen.
Productspecificaties
Specificatie
Details
ProducttypeReagens voor celafhechting / dissociatie
FormaatSteriel gefilterde vloeistof, gebruiksklaar
Inhoud100 ml
Werkconcentratie1x (gebruiksklaar)
EnzymactiviteitGecombineerd proteolytisch en collagenolytisch
EnzymherkomstNiet-zoogdier en niet-bacterieel
BuffersysteemDulbecco’s PBS met EDTA
pH-indicatorFenolrood
pH-bereik6,8 – 7,8
UiterlijkHeldere, lichtrode tot oranje oplossing
Opslagtemperatuur-15 °C of lager
Stabiliteit na ontdooienTot 2 maanden bij +2 °C tot +8 °C
Aanbevolen gebruiksvolume~10 ml per 75 cm² kweekoppervlak
Typische incubatietijd5 – 10 minuten bij kamertemperatuur
VerzendvoorwaardenIngevroren op droogijs
Beoogd gebruikUitsluitend voor onderzoek en verdere productie
Formulering (samenstelling per liter)
Bestanddeel
Concentratie (mg/l)
Anorganische zouten
Natriumchloride (NaCl)8000,00
Dinatriumwaterstoffosfaat (Na2HPO4)1150,00
Kaliumchloride (KCl)200,00
Kaliumdihydrogenofosfaat (KH2PO4)200,00
Overige componenten
EDTA · 4Na (tetranatrium-EDTA)220,00
Fenolrood3,00
Gepatenteerd enzymmengsel (proteolytische en collagenolytische activiteit)1x
Accutase is een geregistreerd handelsmerk van Innovative Cell Technologies, Inc.
, w: 2 mM L-Glutamine, w: 2,2 g/L NaHCO3, w: EBSS")
Deze gebruiksklare, steriel gefilterde vloeibare formulering is aangevuld met Earle’s Balanced Salt Solution (EBSS), 2 mM L-glutamine, D-glucose (1,0 g/l) en 2,2 g/l natriumbicarbonaat (NaHCO3), waardoor het geschikt is voor gebruik in een CO2-geregelde incubatoratmosfeer (doorgaans 5 % CO2). Het toegevoegde fenolrood fungeert als pH-indicator, waardoor de toestand van het medium tijdens de celkweek gemakkelijk visueel kan worden gecontroleerd.
Belangrijkste kenmerken
Klassieke Eagle’s MEM-formulering met Earle’s Balanced Salt Solution (EBSS)
2 mM L-glutamine inbegrepen – klaar voor onmiddellijk gebruik
2,2 g/l natriumbicarbonaat – gebufferd voor incubatie bij 5 % CO2
Met D-glucose (1,0 g/l) als primaire koolstofbron
Met fenolrood als pH-indicator
Zonder HEPES en zonder natriumpyruvaat
Steriel gefilterd vloeibaar medium, klaar voor gebruik
pH 7,0 – 7,6
Typische toepassingen
EMEM ondersteunt de kweek van een breed scala aan zoogdiercellijnen, waaronder HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 en vele primaire cellen. Veelvoorkomende toepassingen zijn onder meer:
Routinematig onderhoud en uitbreiding van hechtende cellijnen
Werkprocessen voor virusvermeerdering en vaccinproductie
Toepassingen op het gebied van cytotoxiciteit en bioassays
Transfectie
- en eiwitexpressiestudies
Fundamenteel onderzoek in celbiologie en moleculaire biologie
Voor optimale celgroei wordt EMEM doorgaans aangevuld met 5–10 % foetaal runderserum (FBS) en, afhankelijk van de cellijn, met niet-essentiële aminozuren (NEAA) en antibiotica zoals penicilline/streptomycine.
Behandeling en opslag
Bewaar de ongeopende fles bij +2 °C tot +8 °C, beschermd tegen licht. Na opening onder aseptische omstandigheden gebruiken. L-glutamine in oplossing is onderhevig aan geleidelijke afbraak – voor de beste prestaties raden wij aan het medium binnen 4 weken na opening te gebruiken, of aan te vullen met verse L-glutamine voor gebruik indien het voor langere tijd wordt bewaard. Laat het medium opwarmen tot 37 °C voordat het aan de cellen wordt toegevoegd.
Kwaliteit
Vervaardigd volgens strenge kwaliteitsnormen. Elke batch wordt getest op steriliteit, pH, osmolaliteit en endotoxineniveaus om consistente prestaties bij celkweektoepassingen te garanderen.
Productspecificaties
Specificatie
Details
ProducttypeMEM
ProductcategorieKweekmedia
FormaatVloeibaar
SterielJa
Inhoud500 ml
L-glutamineMet L-glutamine (2 mM)
GlucoseMet glucose (1,0 g/l)
NatriumwaterstofcarbonaatMet NaHCO3 (2,2 g/L)
HEPESZonder HEPES
Natrium-pyruvaatZonder natriumpyruvaat
FenolroodMet fenolrood
ZoutoplossingEarle’s Balanced Salt Solution (EBSS)
pH7,0 – 7,6
EndotoxinegehalteNiet gespecificeerd
Opslag+2 °C tot +8 °C
Samenstelling (per liter)
Bestanddeel
Concentratie (mg/l)
Anorganische zouten
Calciumchloride · 2H2O265,00
Magnesiumsulfaat97,72
Kaliumchloride400,00
Natriumchloride6.800,00
Natriumdihydrogeenfosfaat, watervrij122,00
Natriumbicarbonaat (NaHCO3)2.200,00
Aminozuren
L-Arginine · HCl126,00
L-cystine · 2HCl31,30
L-Glutamine292,00
L-histidine · HCl · H2O42,00
L-Isoleucine52,00
L-Leucine52,00
L-Lysine · HCl72,50
L-Methionine15,00
L-fenylalanine32,00
L-Threonine48,00
L-tryptofaan10,00
L-Tyrosine · 2Na · 2H2O51,90
L-valine46,00
Vitaminen
D-Calciumpantothenaat1,00
Cholinechloride1,00
Foliumzuur1,00
myo-inositol2,00
Nicotinamide1,00
Pyridoxal · HCl1,00
Riboflavine0,10
Thiamine · HCl1,00
Overige bestanddelen
D(+)-Glucose1.000,00
Fenolrood10,00

De belangrijkste kenmerken van Freeze Medium CM-1 zijn:
Brede compatibiliteit: Effectief voor een breed scala aan celtypen, waaronder primaire cellen, stamcellen en gevestigde cellijnen.
Hoge levensvatbaarheid: Geoptimaliseerd voor maximaal herstel en levensvatbaarheid van de cellen na ontdooiing, voor betrouwbare experimentele resultaten.
Gebruiksklaar: Gemakkelijk bereid en gesteriliseerd voor onmiddellijke toepassing, waardoor de voorbereidingstijd korter wordt en het risico op besmetting kleiner.
Verbeterde stabiliteit: Behoudt consistente prestaties onder standaard cryoconserveringsomstandigheden, waardoor reproduceerbare resultaten worden gegarandeerd.
Lange houdbaarheid: CM-1 is een serumbevattend, gebruiksklaar cryoconserveringsmedium dat tot een jaar in de koelkast kan worden bewaard.
CM-1 gebruiken voor het invriezen van cellen
Volg deze stappen om CM-1 te gebruiken voor het invriezen van zowel adherente als suspensiecellen
Voor adherente cellen, wassen en losmaken van het kweeksubstraat. Voor suspensiecellen gaat u direct door naar de volgende stap.
Tel de cellen om er zeker van te zijn dat ze de juiste concentratie hebben.
Centrifugeer de cellen om ze te pelleteren en resuspendeer ze vervolgens in CM-1 diepvriesmedium.
Breng de geresuspendeerde cellen over in cryovaatjes.
Gebruik een methode voor langzaam invriezen voordat de cellen worden overgebracht naar langdurige opslag
Methode
Beschrijving
Stappen
❄️
Handmatig invriezen
Een stapsgewijze methode waarbij de temperatuur geleidelijk wordt verlaagd om de levensvatbaarheid van de cellen te garanderen
1️⃣ Plaats cellen in vriesmedium in een vriezer van 4°C gedurende 40 minuten.
2️⃣ Breng de cellen over naar een vriezer van -80°C gedurende 24 uur.
3️⃣ Bewaar de cellen in vloeibare stikstof voor langdurige bewaring
❄️
Mr. Frosty gebruiken
Een handig apparaat voor het gecontroleerd invriezen van cellen zonder elektrische voeding
1️⃣ Bereid cellen voor in cryovials met vriesmedium.
2️⃣ Plaats cryovials in Mr. Frosty container.
3️⃣ Bewaar bij -80°C gedurende 24 uur alvorens over te hevelen naar vloeibare stikstof
❄️
Vriezer met gecontroleerde snelheid
Een zeer nauwkeurige vriezer van Thermo Fisher of andere fabrikanten, ontworpen voor gecontroleerde temperatuurverlaging
1️⃣ Programmeer het apparaat om de temperatuur geleidelijk te laten dalen.
2️⃣ Plaats de geprepareerde cellen in de vriezer.
3️⃣ Breng de cellen na de vriescyclus over in vloeibare stikstof
Bewaar de cryovials bij temperaturen lager dan -130°C of in vloeibare stikstof voor langdurige bewaring.
Ingrediënten
Bevat FBS, DMSO, Glucose, Zouten
Buffercapaciteit: pH = 7,2 tot 7,6
Cytion's Freeze Medium CM-1 biedt een betrouwbare oplossing voor cryopreservatie en zorgt voor een hoge levensvatbaarheid en functionaliteit van de cellen na ontdooien voor een breed scala aan onderzoekstoepassingen.

Ham's F-12K (Kaighn's) medium is zorgvuldig geformuleerd om de omstandigheden voor celkweek te optimaliseren. Het heeft een verrijkte samenstelling en biedt verhoogde gehaltes aan essentiële componenten zoals aminozuren en natriumpyruvaat, evenals extra elementen zoals putrescine, thymidine, hypoxanthine en zink. Deze toevoegingen stellen onderzoekers in staat om het medium aan te vullen met minimaal serum of gedefinieerde componenten voor specifieke celtypes, waardoor precieze experimentele omstandigheden mogelijk worden.
Met name Ham's F-12K (Kaighn's) medium bevat geen eiwitten of groeifactoren. Bijgevolg is aanvulling met groeifactoren en foetaal runderserum (FBS) vaak noodzakelijk, waardoor onderzoekers het medium kunnen aanpassen aan de vereisten van hun specifieke cellijnen. Voor optimale prestaties moet de concentratie FBS zorgvuldig worden geoptimaliseerd voor elke cellijn, om optimale groei en functionaliteit te garanderen.
Om de fysiologische pH te handhaven, maakt Ham's F-12K (Kaighn's) medium gebruik van een natriumbicarbonaat buffersysteem (2,5 g/L), waardoor een gecontroleerde 5-10% CO2 omgeving nodig is tijdens de kweek. Dit zorgt ervoor dat de pH van het medium binnen het ideale bereik blijft voor celgroei en levensvatbaarheid
Kwaliteitscontrole
pH = 7,2 +/
- 0,02 bij 20-25°C.
Elk lot is getest op steriliteit en afwezigheid van mycoplasma en bacteriën.
Onderhoud
Koel bewaren bij +2°C tot +8°C in het donker. Invriezen en opwarmen tot +37°C minimaliseren de kwaliteit van het product.
Verwarm het medium niet tot meer dan 37°C en gebruik geen oncontroleerbare warmtebronnen (bijv. magnetronapparatuur).
Als slechts een deel van het medium wordt gebruikt, haal dan deze hoeveelheid uit de fles en warm het op bij kamertemperatuur.
Houdbaarheid voor elk medium behalve het basismedium is 8 weken vanaf de productiedatum.
Samenstelling
Bestanddelen
mg/L
Anorganische zouten
Calciumchloride x 2H2O
135,24
Koper(II)sulfaat x 5H2O
0,00
IJzer(II)sulfaat x 7H2O
0,83
Magnesiumchloride x 6H2O
105,72
Magnesiumsulfaat x 7H2O
394,49
Kaliumchloride
283,29
Kaliumdiwaterstoffosfaat
58,52
Natriumchloride
7597,20
dinatriumwaterstoffosfaatwatervrij
115,02
Zinksulfaat x 7H2O
0,14
Andere componenten
D(+)-Glucose watervrij
1260,00
Hypoxanthine
4,08
DL-α liponzuur
0,21
Fenol rood
3,00
Putrescine x 2HCl
0,32
Natriumpyruvaat
220,00
NaHCO3
2500,00
Thymidine
0,73
Aminozuren
L-Alanine
17,82
L-Arginine x HCl
421,40
L-Asparagine x H2O
30,02
L-Asparaginezuur
26,62
L-Cysteïne x HCl x H2O
70,24
L-Glutamine
292,20
L-Glutaminezuur
29,42
Glycine
15,01
L-Histidine x HCl x H2O
41,92
L-Isoleucine
7,87
L-Leucine
26,24
L-Lysine x HCl
73,04
L-Methionine
8,95
L-Fenylalanine
9,91
L-proline
69,06
L-Serine
21,02
L-Threonine
23,82
L-Tryptofaan
4,08
L-Tyrosine
10,87
L-Valine
23,42
Vitaminen
D(+)-Biotine
0,07
D-Calcium pantothenaat
0,48
Choline chloride
13,96
Foliumzuur
1,32
myo-Inositol
18,02
Nicotinamide
0,04
Pyridoxine x HCl
0,06
Riboflavine
0,04
Thiamine x HCl
0,34
Vitamine B12
1,36

Fosfaatgebufferde zoutoplossing (PBS) is een veelgebruikte bufferoplossing in biologisch en chemisch onderzoek. Het speelt een cruciale rol bij het handhaven van de pH-balans en osmolariteit tijdens verschillende experimentele procedures, waaronder weefselverwerking en celkweek. Onze PBS-oplossing is zorgvuldig geformuleerd met hoogzuivere ingrediënten om stabiliteit en betrouwbaarheid bij elk experiment te garanderen. De osmolariteit en ionenconcentraties van onze PBS benaderen die van het menselijk lichaam, waardoor het isotoon is en niet giftig voor de meeste cellen.
Samenstelling van onze PBS-oplossing
Onze PBS-oplossing is een op pH aangepast mengsel van ultrazuivere fosfaatbuffers en zoutoplossingen. Bij een werkconcentratie van 1X bevat het:
8000 mg/L natriumchloride (NaCl)
200 mg/L Kaliumchloride (KCl)
1150 mg/L Watervrij natriumfosfaat dibasisch (Na2HPO4)
200 mg/L Watervrij monobasisch kaliumfosfaat (KH2PO4)
Deze samenstelling zorgt voor een optimale pH-waarde en ionenbalans, geschikt voor een breed scala aan biologische toepassingen.
Toepassingen van onze PBS-oplossing
Onze PBS-oplossing is ideaal voor verschillende toepassingen in biologisch onderzoek. De isotone en niet-toxische eigenschappen maken het geschikt voor het verdunnen van stoffen en het spoelen van celcontainers. PBS-oplossingen met EDTA zijn effectief voor het losmaken van vastzittende en samengeklonterde cellen. Tweewaardige metalen zoals zink mogen echter niet aan PBS worden toegevoegd, omdat dit tot precipitatie kan leiden. In dergelijke gevallen worden Good's buffers aanbevolen. Bovendien is onze PBS-oplossing een aanvaardbaar alternatief voor virale transportmedia voor het transport en de opslag van RNA-virussen, waaronder SARS-CoV-2.
Kwaliteitscontrole
Steriel gefilterd
Opslag en houdbaarheid
Bewaren bij +2°C tot +25°C, beschermd tegen licht.
Eenmaal geopend, bewaren bij 2°C tot 25°C en binnen 24 maanden gebruiken.
Verzendomstandigheden
Omgevingstemperatuur
Onderhoud
Koel bewaren bij +2°C tot +8°C in het donker. Vermijd invriezen en frequente opwarming tot +37°C, omdat dit de kwaliteit van het product vermindert.
Verwarm het medium niet verder dan 37°C en gebruik geen ongecontroleerde warmtebronnen zoals magnetrons.
Als slechts een deel van het medium gebruikt wordt, verwijder dan de benodigde hoeveelheid en warm het op tot kamertemperatuur voor gebruik.
Samenstelling
Categorie
Bestanddelen
Concentratie (mg/L)
Zouten
Kaliumchloride
200
Kaliumfosfaat monobasisch watervrij
200
Natriumchloride
8000
Watervrij natriumfosfaat dibasisch
1150

In eerste instantie ontworpen om de groei van menselijke leukemische cellen te ondersteunen in zowel suspensie
- als monolaagculturen, heeft RPMI 1640 Medium zich ontwikkeld door aanpassingen door onderzoekers en commerciële leveranciers om geschikt te worden voor een breed scala aan zoogdiercellen. Het is uitzonderlijk compatibel met cellijnen zoals HeLa, Jurkat, MCF-7, PC12, PBMC, astrocyten en carcinomen.
RPMI 1640 Medium onderscheidt zich van andere celkweekmedia door zijn unieke samenstelling. Het bevat een aanzienlijke hoeveelheid fosfaat, aminozuren en vitaminen. Het bevat met name biotine, vitamine B12 en PABA, die afwezig zijn in Eagle's Minimal Essential Medium of Dulbecco's Modified Eagle Medium. Bovendien vertoont RPMI 1640 Medium significant verhoogde concentraties vitamine inositol en choline. Het bevat echter geen eiwitten, lipiden of groeifactoren. Daarom is aanvulling met 10% foetaal runderserum (FBS) vaak nodig om optimale omstandigheden voor celgroei te creëren.
Het buffersysteem van RPMI 1640 Medium is gebaseerd op natriumbicarbonaat en vereist een omgeving met 5-10% CO2 om een fysiologisch geschikte pH te handhaven. De toevoeging van het reductiemiddel glutathion onderscheidt dit medium verder van andere.
Kwaliteitscontrole
Steriel gefilterd
Opslag en houdbaarheid
Bewaren bij +2°C tot +8°C, beschermd tegen licht.
Na opening bewaren bij 4 °C en binnen 6-8 weken gebruiken.
Verzendomstandigheden
Omgevingstemperatuur
Onderhoud
Koel bewaren bij +2°C tot +8°C in het donker. Vermijd invriezen en frequente opwarming tot +37°C, omdat dit de kwaliteit van het product vermindert.
Verwarm het medium niet verder dan 37°C en gebruik geen ongecontroleerde warmtebronnen zoals magnetrons.
Als slechts een deel van het medium gebruikt wordt, verwijder dan de benodigde hoeveelheid en warm het op tot kamertemperatuur voor gebruik.
Samenstelling
Categorie
Bestanddelen
Concentratie (mg/L)
Aminozuren
Glycine
10.00
L-Alanyl-L-Glutamine
434.40
L-Arginine
200.00
L-AsparagineH2O
56.82
L-Asparaginezuur
20.00
L-Cystine 2HCl
65.20
L-Glutaminezuur
20.00
L-Histidine HClH2O
20.27
L-Hydroxy-L-proline
20.00
L-Isoleucine
50.00
L-Leucine
50.00
L-Lysine HCl
40.00
L-Methionine
15.00
L-Fenylalanine
15.00
L-Proline
20.00
L-Serine
30.00
L-Threonine
20.00
L-Tryptofaan
5.00
L-Tyrosine 2Na 2H2O
28.83
L-Valine
20.00
Vitaminen
p-Amino Benzoëzuur
1.00
D-Biotine
0.20
Choline Chloride
3.00
D-Calcium Pantothenaat
0.25
Foliumzuur
1.00
myo-Inositol
35.00
Nicotinamide
1.00
Pyridoxine HCl
1.00
Riboflavine
0.20
Thiamine HCl
1.00
Vitamine B12
0.005
Anorganische zouten
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Andere componenten
D-glucose
2000.00
L-Glutathion Verminderd
1.00
Fenolrood natriumzout
5.30
, w: 3,1 g/L Glucose, w: 2,5 mM L-Glutamine, w: 15 mM HEPES, w: 0,5 mM Natriumpyruvaat, w: 1,2 g/L NaHCO3")
Deze unieke formulering combineert Dulbecco's Modified Eagle Medium (DMEM) en Ham's F-12 (Ham's Nutrient Mixture F-12) in een precieze verhouding van 1:1. De toevoeging van L-glutamine verbetert de samenstelling nog verder.
DMEM, afgeleid van Eagle's Minimal Essential Medium (EMEM), biedt een hogere concentratie aan aminozuren en vitamines in vergelijking met zijn voorganger. Ham's F-12 daarentegen is gebaseerd op Ham's F-10-medium en biedt een aanvullende reeks essentiële componenten.
Om een optimale celgroei te ondersteunen, is het gebruikelijk om DMEM:Ham's F12 aan te vullen met FBS in een concentratie van doorgaans 5-10%. Deze toevoeging is noodzakelijk omdat het medium geen groeihormonen, lipiden en eiwitten bevat die cruciaal zijn voor de celontwikkeling.
DMEM:Ham's F12 bevat een pH-buffersysteem en wordt vaak aangevuld met fenolrood, een pH-indicator. Gekweekte cellen in DMEM:Ham's F12, of elk ander medium dat gebruikmaakt van het bicarbonaatbuffersysteem, vereisen een gecontroleerde CO2-omgeving van 5-10% om de juiste pH-waarden te handhaven.
Kwaliteitscontrole
Steriel gefilterd
Opslag en houdbaarheid
Bewaren bij +2 °C tot +8 °C, beschermd tegen licht.
Na opening bewaren bij 4 °C en binnen 6–8 weken gebruiken.
Verzendvoorwaarden
Omgevingstemperatuur
Bewaring
Bewaar gekoeld bij +2 °C tot +8 °C in het donker. Vermijd bevriezing en herhaaldelijk opwarmen tot +37 °C, aangezien dit de productkwaliteit vermindert.
Verwarm het medium niet boven 37 °C en gebruik geen ongecontroleerde warmtebronnen zoals magnetrons.
Als slechts een deel van het medium wordt gebruikt, neem dan de benodigde hoeveelheid en verwarm deze tot kamertemperatuur voor gebruik.
Samenstelling
Categorie
Bestanddelen
Concentratie (mg/L)
Aminozuren
Glycine
18,75
L-alanine
4,45
L-Arginine HCl
147,50
L-asparagine H₂O
7,50
L-asparaginezuur
6,65
L-cysteïne HCl H₂O
17,56
L-Cystine 2 HCl
31,29
L-glutaminezuur
7,35
L-glutamine
365,00
L-histidine HCl H₂O
31,48
L-Isoleucine
54,47
L-Leucine
59,05
L-Lysine HCl
91,25
L-methionine
17,24
L-fenylalanine
35,48
L-proline
17,25
L-serine
26,25
L-Threonine
53,45
L-tryptofaan
9,02
L-Tyrosine 2 Na 2 H2O
55,79
L-valine
52,85
Vitaminen
D-Biotine
0,0035
Cholinechloride
8,98
D-Calciumpantothenaat
2,24
Foliumzuur
2,66
myo-inositol
12,60
Nicotinamide
2,02
Pyridoxine HCl
0,031
Pyridoxal HCl
2,00
Riboflavine
0,219
Thiamine HCl
2,17
Vitamine B12
0,68
Anorganische zouten
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO₃)₃ 9 H₂O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Overige componenten
D-glucose
3151,00
Hypoxanthine
2,40
HEPES
3574,50
Linolzuur
0,042
Liponzuur
0,105
Fenolrood-natriumzout
8,63
Putrescine-2-HCl
0,081
Natrium-pyruvaat
55,00
Thymidine
0,365
, w: 3,0 g/L Glucose, w: stabiel Glutamine, w: 2,0 mM Natriumpyruvaat, w: 2,2 g/L NaHCO3")
- 0,02 bij 20-25 °C. Elke partij is getest op steriliteit en de afwezigheid van mycoplasma en bacteriën. Bewaring Bewaar gekoeld bij +2 °C tot +8 °C in het donker. Invriezen en opwarmen tot +37 °C vermindert de kwaliteit van het product. Verwarm het medium niet tot meer dan 37 °C en gebruik geen oncontroleerbare warmtebronnen (bijv. magnetrons). Als slechts een deel van het medium wordt gebruikt, haal dan deze hoeveelheid uit de fles en verwarm deze op kamertemperatuur. De houdbaarheid van alle media, met uitzondering van het basismedium, is 6 tot 8 weken vanaf de datum van opening. Samenstelling Bestanddelen mg/L Anorganische zoutenCalciumchloride x 2H2O132,00 Magnesiumsulfaat97,67 Kaliumchloride400,00 Natriumchloride6.460,00 Dinatriumwaterstoffosfaat (watervrij)504,00 Overige componentenD(+)-glucose (watervrij)3.000,00 Glutathion (gereduceerd)0,50 Vleespeptone600,00 Fenolrood natriumzout11,00 AminozurenL-alanine13,36 L-arginine x HCl42,14 L-asparagine x H2O45,03 L-asparaginezuur19,97 L-Cysteïne x HCl x H2O31,75 L-glutamine (stabiel)219,15 L-glutaminezuur22,07 Glycine7,51 L-Histidine x HCl x H2O20,96 L-hydroxyproline19,67 L-Isoleucine39,36 L-Leucine39,36 L-lysine x HCl36,54 L-methionine14,92 L-fenylalanine16,52 L-proline17,27 L-serine26,28 L-threonine17,87 L-tryptofaan3,06 L-Tyrosine dinatriumbrood26,10 L-valine17,57 Vitaminenp-Aminobenzoëzuur1,00 Ascorbinezuur0,56 D(+)-biotine0,20 D-calciumpantothenaat0,20 Cholinechloride5,00 Foliumzuur10,0 Myo-inositol36,00 Nicotinamide0,50 Nicotinezuur0,50 Pyridoxal HCl0,50 Pyridoxine HCl0,50 Riboflavine0,20 Thiamine HCl0,20 Vitamine B122,00

Medium 199 biedt een scala aan toepassingen in het veld. Het kan het cumulus-oocytcomplex (COC) effectief in stand houden en de in vitro rijping van oöcyten ondersteunen. Daarnaast wordt het gebruikt bij het spoelen van aspiratielijnen tijdens het verzamelen van eicellen van Duitse Holstein-koeien. Bovendien dient Medium 199 als een uitstekend medium voor de kweek van cardiale endotheelcellen afkomstig van ratten. Deze toepassingen tonen de veelzijdigheid en aanpasbaarheid van Medium 199 aan verschillende experimentele behoeften.
Geschiedenis
De ontwikkeling van medium 199 in de jaren 1950 betekende een belangrijke vooruitgang op het gebied van weefselkweekmedia. Vóór de introductie waren veel kweekmedia afhankelijk van dierlijke producten en weefselextracten. Morgan en collega's brachten echter een revolutie teweeg door een volledig gedefinieerde voedingsbron voor celculturen te formuleren. Door hun experimenten met verschillende combinaties van vitamines, aminozuren en andere factoren ontdekten ze de uitzonderlijke groeibevorderende eigenschappen van Medium 199.
Kwaliteitscontrole
pH = 7,2 +/
- 0,02 bij 20-25°C.
Elke partij is getest op steriliteit en afwezigheid van mycoplasma en bacteriën.
Onderhoud
Koel bewaren bij +2°C tot +8°C in het donker. Invriezen en opwarmen tot +37°C minimaliseren de kwaliteit van het product.
Verwarm het medium niet tot meer dan 37° C en gebruik geen oncontroleerbare warmtebronnen (bijv. magnetronapparatuur).
Als slechts een deel van het medium wordt gebruikt, haal dan deze hoeveelheid uit de fles en warm het op bij kamertemperatuur.
Houdbaarheid voor elk medium behalve het basismedium is 8 weken vanaf de productiedatum.
Samenstelling
Bestanddelen
mg/L
Anorganische zouten
Calciumchloride x 2H2O
264,92
IJzer(III)nitraat x 9H2O
0,72
Magnesiumsulfaat
97,67
Kaliumchloride
400,00
Natriumacetaat x 3H2O
82,95
Natriumchloride
6,800.00
Natriumdiwaterstoffosfaat x H2O
140,00
Andere componenten
Adenine sulfaat
10,00
AMP
0,20
ATP
1,00
Cholesterol
0,20
2'-Deoxyribose
0,50
D(+)-Glucose watervrij
1,000.00
Glutathion (rood.)
0,05
Guanine x HCl
0,30
Hypoxanthine
0,30
Fenol rood
10,00
D-Ribose
0,50
Thymine
0,30
Tween 80
4,90
Uracil
0,30
Xanthine
0,30
NaHCO3
2,200.00
Aminozuren
L-Alanine
25,00
L-Arginine x HCl
70,00
L-Asparaginezuur
30,00
L-Cysteïne x HCl x H2O
0,10
L-Cystine
20,00
L-Glutamine stabiel
149,00
L-Glutaminezuur
67,00
Glycine
50,00
L-Histidine x HCl x H2O
21,88
L-Hydroxyproline
10,00
L-Isoleucine
20,00
L-Leucine
60,00
L-Lysine x HCl
70,00
L-Methionine
15,00
L-Fenylalanine
25,00
L-Proline
40,00
L-Serine
25,00
L-Threonine
30,00
L-Tryptofaan
10,00
L-Tyrosine
40,00
L-Valine
25,00
Vitaminen
4-Amino benzoëzuur
0,05
Ascorbinezuur
0,05
D(+)-Biotine
0,01
Calciferol
0,10
D-Calcium pantothenaat
0,01
Choline chloride
0,50
Foliumzuur
0,01
myo-Inositol
0,05
Menadion
0,01
Nicotinezuur
0.025
Nicotinamide
0.025
Pyridoxaal x HCl
0.025
Pyridoxol x HCl
0.025
Riboflavine
0,01
DL-α-tocoferolfosfaat dinatriumzout
0,01
Thiamine x HCl
0,01
Vitamine A-acetaat
0,14

IMDM is zeer geschikt voor snel groeiende celculturen met een hoge dichtheid, waaronder Jurkat-, COS-7
- en macrofaagcellen. De verschillende modificaties van IMDM die beschikbaar zijn voor een reeks celkweektoepassingen kunnen worden gevonden met behulp van de media selector tool. Vloeibare media leveren essentiële voedingsstoffen voor alle celkweektoepassingen. Elk van onze hoogwaardige celkweekmedia wordt geproduceerd volgens de oorspronkelijk gepubliceerde formule of modificaties die nodig zijn voor consistente prestaties en stabiliteit van het kweekmedium.
IMDM vs. DMEM
IMDM bevat kaliumnitraat in plaats van ijzer-nitraat en HEPES en natriumpyruvaat. De extra componenten in IMDM maken het geschikter voor gespecialiseerde celtypes en specifieke toepassingen dan DMEM.
IMDM vs. RPMI
IMDM en RPMI hebben verschillende formuleringen die relevant kunnen zijn voor PMA/ionomycine stimulatie. Een belangrijk verschil is de concentratie Ca2+. Terwijl RPMI 0,42 mM Ca2+ bevat, bevat IMDM 1,49 mM.
Kwaliteitscontrole
pH = 7,2 +/
- 0,02 bij 20-25°C.
Elk lot is getest op steriliteit en afwezigheid van mycoplasma en bacteriën.
Onderhoud
Gekoeld bewaren bij +2°C tot +8°C in het donker. Invriezen en opwarmen tot +37°C minimaliseren de kwaliteit van het product.
Verwarm het medium niet tot meer dan 37° C en gebruik geen oncontroleerbare warmtebronnen (bijv. magnetronapparatuur).
Als slechts een deel van het medium wordt gebruikt, haal dan deze hoeveelheid uit de fles en warm het op bij kamertemperatuur.
Houdbaarheid voor elk medium, behalve het basismedium, is 8 weken vanaf de productiedatum.
Samenstelling
Bestanddelen
mg/L
Anorganische zouten
Calciumchloride x 2 H2O
219,00
Kaliumchloride
330,00
Kaliumnitraat
0.076
Watervrij magnesiumsulfaat
97,73
Natriumchloride
4,505.00
Natriumdiwaterstoffosfaat watervrij
109,00
Natriumseleniet
0,02
Andere componenten
D(+)-Glucose watervrij
4,500.00
HEPES
5,958.00
Natriumpyruvaat
110,00
Fenol rood
15,00
Aminozuren
L-Alanine
25,00
L-Arginine x HCl
84,00
L-Asparagine x H2O
25,00
L-Asparaginezuur
30,00
L-Cystine x 2HCl
91,24
L-Glutamine
584,00
L-Glutaminezuur
75,00
Glycine
30,00
L-Histidine x HCl x H2O
42,00
L-Isoleucine
104,80
L-Leucine
104,80
L-Lysine x HCl
146,20
L-Methionine
30,00
L-Fenylalanine
66,00
L-Proline
40,00
L-Serine
42,00
L-Threonine
95,20
L-Tryptofaan
16,00
L-Tyrosine x 2Na
104,20
L-Valine
93,60
Vitaminen
D(+)-Biotine
0.013
D-Calcium pantothenaat
4,00
Choline chloride
4,00
Foliumzuur
4,00
myo-Inositol
7,20
Celkweekmedia: een overzicht
Op het gebied van de levenswetenschappen is celkweek een van de belangrijkste methoden. Onder de term 'celkweek' wordt verstaan: het verwijderen van cellen, weefsels of organen uit een dier of plant en het vervolgens implanteren van die cellen, weefsels of organen in een kunstmatige omgeving die gunstig is voor hun overleving en/of groei. De fundamentele omgevingsvereisten voor een optimale celontwikkeling zijn een geregelde temperatuur, een substraat waaraan de cellen zich kunnen hechten, een geschikt groeimedium en een incubator die de optimale pH-waarde en osmolaliteit handhaaft. Cellen hebben deze omstandigheden nodig om zich optimaal te kunnen ontwikkelen.
De keuze van een geschikt groeimedium voor in-vitro-kweek is de fase in de celkweek die zowel het meest cruciaal als het meest essentieel is. Een groeimedium, ook wel kweekmedium genoemd, is een vloeistof of gel die is samengesteld om de ontwikkeling van organismen op microscopisch, cellulair of plantachtig niveau te bevorderen. Het medium dat wordt gebruikt voor het kweken van cellen bevat vaak een voldoende voorraad energie en stoffen die de celcyclus regelen. De belangrijkste bestanddelen van een kweekmedium zijn onder meer aminozuren, vitamines, anorganische zouten, glucose en serum. Het serum wordt aan het medium toegevoegd omdat het fungeert als bron van groeifactoren, hormonen en hechtingsfactoren. Naast het leveren van voedingsstoffen draagt het medium ook bij aan het handhaven van de pH- en osmolaliteitsniveaus.
Soorten media die worden gebruikt in celcultuur
Zowel menselijke als dierlijke cellen kunnen worden gekweekt in een kunstmatig of synthetisch medium, of in een volledig natuurlijk medium dat is aangevuld met natuurlijke elementen. Hieronder geven we u een overzicht van de verschillende momenteel beschikbare mediatypen.
Natuurlijke media
In natuurlijke media komen uitsluitend biologische vloeistoffen voor die in hun natuurlijke staat voorkomen. Natuurlijke media zijn zeer geschikt en eenvoudig te gebruiken voor de kweek van een breed scala aan dierlijke celtypen. Het gebrek aan inzicht in de precieze samenstelling van natuurlijke media is de belangrijkste factor die bijdraagt aan de lage herhaalbaarheid van resultaten die met natuurlijke media worden verkregen.
Kunstmatige media
Bij de bereiding van kunstmatige of synthetische media worden voedingsstoffen (zowel organische als anorganische), serumproteïnen, koolhydraten, cofactoren, vitamines en zouten toegevoegd, evenals O₂- en CO₂-gasfasen [1].
Er zijn verschillende soorten kunstmatige media ontwikkeld om aan een of meer van de volgende functies te voldoen: 1) Onmiddellijke overleving (een uitgebalanceerde zoutoplossing met een nauwkeurige pH-waarde en osmotische druk). 2) Langdurige overleving (een uitgebalanceerde zoutoplossing aangevuld met verschillende samenstellingen van organische chemicaliën en/of serum). 3) Onbeperkte ontwikkeling. 4) Gespecialiseerde functies.
Er zijn vier verschillende classificaties voor kunstmatige media:
Media die serum bevatten
Het meest voorkomende supplement in media voor de kweek van dierlijke cellen is foetaal runderserum. Dit wordt als goedkoop supplement aan het kweekmedium toegevoegd om de best mogelijke groeiomstandigheden te bereiken. Het serum fungeert niet alleen als drager of chelaatvormer voor voedingsstoffen die onstabiel of niet in water oplosbaar zijn, hormonen en groeifactoren, proteaseremmers en andere stoffen, maar bindt en neutraliseert ook schadelijke moleculen.
Serumvrij medium
De aanwezigheid van serum in het medium heeft een aantal nadelen en kan leiden tot ernstige interpretatiefouten in immunologisch onderzoek [2, 3]. Er zijn diverse serumvrije media ontwikkeld [4, 5]. Deze media zijn over het algemeen specifiek samengesteld om de kweek van één bepaald celtype te ondersteunen, zoals Knockout Serum Replacement en Knockout DMEM van Thermo Fisher Scientific, en mTESR-medium van Stem Cell Technologies [6], voor stamcellen [7].
Bovendien bevatten deze media vastgestelde hoeveelheden gezuiverde groeifactoren, lipoproteïnen en andere eiwitten, die anders doorgaans door het serum worden geleverd [8]. Deze media worden vaak „gedefinieerde kweekmedia“ genoemd, aangezien de componenten waaruit deze media bestaan goed bekend zijn.
Chemisch gedefinieerde media
Deze media bevatten ultrazuivere anorganische en organische componenten die niet door enige vorm van verontreiniging zijn aangetast. Ze kunnen ook toevoegingen van zuivere eiwitten bevatten, zoals groeifactoren.
De genetische modificatie van bacteriën of gist, in combinatie met de toevoeging van bepaalde vetzuren, vitamines, cholesterol en aminozuren, leidt tot de productie van hun samenstellende delen [9].
Eiwitvrije media
Eiwitvrije media zijn media die helemaal geen eiwitten bevatten, maar uitsluitend niet-eiwitelementen. In vergelijking met media waaraan serum is toegevoegd, bevordert het gebruik van media zonder toegevoegde eiwitten een grotere celproliferatie en eiwitexpressie en maakt het het zuiveren van producten die in een downstream-proces worden gegenereerd eenvoudiger [10-12]. Eiwit is niet opgenomen in formuleringen zoals MEM en RPMI-1640. Indien nodig kan echter een eiwitsupplement worden toegediend.
Kweekmedia en hun basiscomponenten
Commercieel kweekmedium is verkrijgbaar in poeder- of vloeibare vorm en bevat vaak diverse voedingsstoffen, zoals aminozuren, glucose, zouten, vitamines en andere voedingssupplementen.
De behoefte aan deze componenten verschilt per cellijn, en deze variaties zijn verantwoordelijk voor het grote aantal verschillende samenstellingen van media. Elke component vervult een bepaalde functie, die in de volgende paragrafen wordt beschreven:
Buffersystemen
Om optimale groeiomstandigheden te handhaven, moet de pH worden geregeld, wat vaak gebeurt door middel van een van de twee volgende buffersystemen:
Natuurlijk buffersysteem
De CO₂/H₂CO₃-verhouding in de atmosfeer is gelijk aan die van het medium, waardoor een natuurlijk buffermechanisme ontstaat. Om dit natuurlijke buffermechanisme te behouden, moeten culturen worden gehouden in een luchtomgeving met 5-10% CO₂, wat vaak wordt bereikt door gebruik te maken van een CO₂-incubator. Een van de grootste voordelen van het gebruik van een natuurlijke buffer is dat deze goedkoop en veilig is.
HEPES
Chemische buffering met behulp van het zwitterion HEPES heeft een groter bufferend vermogen in het pH-bereik van 7,2–7,4 en vereist geen gereguleerde gasomgeving. Voor bepaalde celtypen kan een hogere dosis HEPES schadelijk zijn. Media die HEPES bevatten, zijn eveneens veel gevoeliger voor de fototoxische effecten van fluorescentielicht [13].
Fenolrood
De pH-indicator fenolrood wordt vaak toegevoegd aan in de handel verkrijgbare kweekmedia, waardoor de pH continu kan worden gecontroleerd. Naarmate de cellen zich vermenigvuldigen, veroorzaken de door deze cellen geproduceerde metabolieten een verschuiving in de pH en daarmee een kleurverandering van het medium. Fenolrood heeft een dubbel effect op de kleur van het medium: bij een zure pH kleurt het geel en bij een basische pH paars. Bij een pH van 7,4 – de optimale waarde voor celkweek – krijgt het medium een fluorescerende rode kleur.
Maar fenolrood heeft enkele nadelen: ten eerste kan fenolrood de werking van een aantal steroïde hormonen nabootsen, met name oestrogeen [14]. Daarom wordt bij het bestuderen van oestrogeengevoelige cellen, zoals borstweefsel, een medium zonder fenolrood aanbevolen. De natrium-kaliumbalans wordt in verschillende serumvrije formuleringen verstoord door de aanwezigheid van fenolrood. Het toevoegen van serum of runderhypofysehormoon aan de media kan dit effect tegengaan [15]. Ten derde wordt de detectie bij flowcytometrische experimenten belemmerd door de aanwezigheid van fenolrood.
Anorganische zouten
Media die anorganische zouten bevatten, zoals natrium-, kalium- en calciumionen, helpen bij het handhaven van het osmotisch evenwicht en het reguleren van de membraanpotentiaal.
Aminozuren
Aangezien aminozuren de fundamentele bouwstenen van eiwitten zijn, vormen ze een essentieel onderdeel van elk celgroeimedium dat ooit is ontwikkeld. Omdat cellen bepaalde aminozuren niet zelf kunnen aanmaken, is het belangrijk dat het kweekmedium essentiële aminozuren bevat. Ze zijn noodzakelijk voor de celproliferatie, en de concentratie waarin ze aanwezig zijn, bepaalt de maximale celdichtheid die kan worden bereikt. Met name L-glutamine, een essentieel aminozuur, is van cruciaal belang.
L-glutamine fungeert als een secundaire energiebron voor het metabolisme en levert stikstof voor de productie van NAD, NADPH en nucleotiden. Aangezien L-glutamine een onstabiel aminozuur is dat na verloop van tijd verandert in een vorm die cellen niet kunnen benutten, moet het aan het medium worden toegevoegd.
Daarnaast kunnen niet-essentiële aminozuren aan het medium worden toegevoegd om de aminozuren aan te vullen die tijdens het groeiproces zijn verbruikt. De groei van de cellen wordt gestimuleerd en hun levensvatbaarheid neemt toe wanneer het groeimedium wordt aangevuld met niet-essentiële aminozuren.
Koolhydraten
Koolhydraten in de vorm van suikers vormen de belangrijkste energiebron. Veel media bevatten naast de meer gangbare suikers glucose en galactose ook maltose en fructose.
Eiwitten en peptiden
Albumine, transferrine en fibronectine zijn de meest gebruikte eiwitten en peptiden. Ze zijn vooral van belang in media die geen serum bevatten. Albumine, transferrine, aprotinine, fetuine en fibronectine zijn enkele van de eiwitten die in serum kunnen worden aangetroffen, dat een rijke bron van eiwitten is.
Albumine is het belangrijkste eiwit in het bloed en heeft als functie het binden en transporteren van diverse stoffen, waaronder water, zouten, vrije vetzuren, hormonen en vitamines, tussen verschillende organen en cellen. Het vermogen van albumine om zich aan chemische stoffen te binden, maakt het een geschikte kandidaat voor het verwijderen van schadelijke verbindingen uit het medium waarin cellen worden gekweekt.
Aprotinine is een beschermend middel in celkweeksystemen, aangezien het stabiel is bij een neutrale en zure pH, en bovendien bestand is tegen hoge temperaturen en de afbraak die kan worden veroorzaakt door proteolytische enzymen. Het is in staat om een aantal serineproteasen te remmen, waaronder onder andere trypsine.
Fetuïne is een glycoproteïne dat in hogere concentraties kan worden aangetroffen in het serum van foetale en pasgeboren dieren in vergelijking met serum van volwassen dieren. Daarnaast fungeert het als een serineproteaseremmer. Het eiwit fibronectine is een essentieel onderdeel van het proces van celadhesie. Transferrine is een eiwit dat ijzer transporteert en verantwoordelijk is voor de afgifte van ijzer aan de celmembranen.
Vetzuren en lipiden
Ze spelen een cruciale rol in serumvrij medium wanneer er geen serum aanwezig is.
Vitaminen
Talrijke vitamines zijn noodzakelijk voor de celontwikkeling en -proliferatie. Vitamines kunnen niet in voldoende hoeveelheden door cellen worden aangemaakt en zijn daarom in weefselcultuur onmisbaar als voedingssupplementen.
In celcultuur is serum de belangrijkste bron van vitamines; media worden echter ook verrijkt met diverse vitamines om ze geschikt te maken voor een specifiek celtype. Meestal worden de vitamines uit de B-groep gebruikt om de celgroei te stimuleren.
Sporenelementen
Chemische elementen zoals koper, zink, selenium en tricarbonzuur-tussenproducten staan bekend als sporenelementen. Sporenelementen worden vaak toegevoegd aan media die geen serum bevatten, ter vervanging van de elementen die doorgaans in serum aanwezig zijn. Deze elementen zijn belangrijke chemische componenten die nodig zijn voor een gezonde celontwikkeling. Veel biochemische reacties zijn afhankelijk van bepaalde micronutriënten, zoals enzymactiviteit.
Mediumsupplementen
Het volledige groeimedium dat voor bepaalde cellijnen wordt aanbevolen, heeft extra componenten nodig die ontbreken in de basismedia en het serum. Deze voedingssupplementen ondersteunen de celgroei en een goede stofwisseling.
Hoewel hormonen, groeifactoren en signaalmoleculen essentieel zijn voor de juiste proliferatie van bepaalde cellijnen, moeten altijd de volgende voorzorgsmaatregelen worden genomen: Aangezien de toevoeging van supplementen de osmolaliteit van het volledige groeimedium kan veranderen, wat de celontwikkeling kan remmen, is het altijd raadzaam om de osmolaliteit te controleren na het toevoegen van supplementen. Voor de meeste cellijnen ligt de optimale osmolaliteit tussen 260 en 320 mOSM/kg.
Antibiotica
Antibiotica worden vaak ingezet om de groei van bacteriële en schimmelverontreinigingen te remmen [16], hoewel ze niet essentieel zijn voor de celgroei. Aangezien antibiotica besmetting met mycoplasma en resistente bacteriën kunnen maskeren, wordt routinematig gebruik ervan bij celculturen afgeraden [17, 18].
Bovendien kunnen antibiotica het metabolisme van overgevoelige cellen verstoren. De penicilline-streptomycine-combinaties van MilliporeSigma en Life Technologies worden vaak gebruikt. Plasmocin is gebruikt bij de kweek van de glioomcellijnen TS603, TS516 en BT260 [19], en is effectief gebleken bij het verwijderen van mycoplasma-besmetting (20).
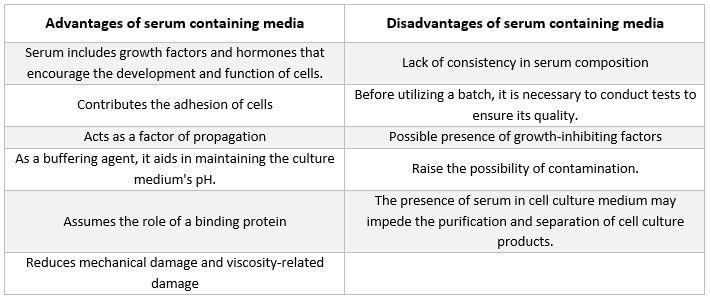
Serum
Albumine, groeifactoren en groeiremmers zijn allemaal aanwezig in serum. Serum is een van de belangrijkste componenten van celkweekmedium omdat het aminozuren, eiwitten, vitamines (met name in vet oplosbare vitamines zoals A, D, E en K), koolhydraten, lipiden, hormonen, groeifactoren, mineralen en sporenelementen levert.
Serum afkomstig van foetussen en kalveren wordt vaak gebruikt om de ontwikkeling van gekweekte cellen te bevorderen. Foetaal serum is een rijke bron van groeifactoren en is geschikt voor het klonen van cellen en de ontwikkeling van gevoelige cellen. Vanwege zijn verminderde groeibevorderende eigenschappen wordt kalfsserum gebruikt in contactinhibitie-experimenten. Normale kweekmedia bevatten vaak 2% tot 10% serum. De toevoeging van serum aan kweekmedium dient de volgende doelen [21]:
-
Het serum levert de essentiële voedingsstoffen voor cellen (zowel in oplossing als gebonden aan eiwitten).
-
Serum bevat verschillende groeifactoren en hormonen die betrokken zijn bij groeibevordering en gespecialiseerde celactiviteit.
-
Het biedt talrijke bindende eiwitten, zoals albumine en transferrine, die andere stoffen naar de cel transporteren. Albumine brengt bijvoorbeeld vetten, vitamines, hormonen enz. naar de cellen.
-
Het levert ook eiwitten, zoals fibronectine, die de celhechting aan het substraat vergroten. Daarnaast produceert het spreidingselementen die helpen bij de celuitbreiding vóór de deling.
-
Het levert proteaseremmers die proteolyse in cellen voorkomen.
-
Het bevat ook mineralen zoals Na+, K+, Zn2+ en Fe2+.
-
Het verhoogt de viscositeit van het medium, waardoor de cellen worden beschermd tegen mechanische beschadiging tijdens het roeren van de suspensiecultuur.
-
Het fungeert tevens als buffer.
Referenties
[1] Morgan J, Morton H, Parker R. Voeding van dierlijke cellen in weefselkweek; eerste onderzoeken naar een synthetisch medium. Proc Soc Exp Biol Med. 1950;73:1-8
[2] Kerbel R, Blakeslee D. Snelle adsorptie van een component van foetaal kalfsserum door zoogdiercellen in kweek. Een mogelijke bron van artefacten in onderzoeken naar antiserum tegen celspecifieke antigenen. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. Toevoeging van serum aan het medium dat wordt gebruikt voor de bereiding van celsuspensies als mogelijke bron van artefacten bij celgemedieerde reacties, onderzocht met behulp van de popliteale lymfeklierproef. J Immunogenet. 1980;7:483-9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Commerciële serumvrije media: groei van hybridomen en productie van monoklonale antilichamen. J Immunol Methods. 1991;145:175-83
[5] Barnes D, Sato G. Methoden voor de kweek van gekweekte cellen in serumvrij medium. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O, et al. HSP70 begeleidt RNA-vrij TDP-43 naar anisotrope intranucleaire vloeibare bolvormige schillen. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, et al. Door het Down-syndroom geïnduceerde senescentie verstoort de nucleaire architectuur van neurale voorlopercellen. Cell Stem Cell. 2022;29:116-130.e7
[8] Iscove N, Melchers F. Volledige vervanging van serum door albumine, transferrine en sojalipiden in culturen van lipopolysaccharide-reactieve B-lymfocyten. J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Systematische verbetering van een chemisch gedefinieerd eiwitvrij medium voor de groei van hybridomen en de productie van monoklonale antilichamen. J Biotechnol. 1996;45:111-23
[10] Darfler F. Een eiwitvrij medium voor de groei van hybridomen en andere cellen van het immuunsysteem. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Serumvrije celkweek: een uniforme aanpak. Cell. 1980;22:649-55
[12] Hamilton W, Ham R. Klonale groei van Chinese hamstercellijnen in eiwitvrije media. In Vitro. 1977;13:537-47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Analyse van de cytotoxische effecten van aan licht blootgesteld HEPES-bevattend kweekmedium. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. Fenolrood in weefselkweekmedia is een zwak oestrogeen: implicaties voor het onderzoek naar oestrogeen-responsieve cellen in kweek. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Ontwikkeling van serumvrije media. In: Master JRW, red. Animal Cell culture, 3e druk. Oxford: Oxford University Press; 2000.
[16] Perlman D. Het gebruik van antibiotica in celkweekmedia. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. Verspreiding en bestrijding van mycoplasma-infecties in celculturen. In Vitro. 1976;12:643-8
[18] Masters J, Stacey G. Het verversen van medium en het passeren van cellijnen. Nat Protoc. 2007;2:2276-84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M, et al. Histon-demethylase KDM6A detecteert zuurstof direct om het chromatine en het lot van de cel te regelen. Science. 2019;363:1217-1222
[20] Molla Kazemiha V, Azari S, Amanzadeh A, Bonakdar S, Shojaei Moghadam M, Habibi Anbouhi M, et al. De werkzaamheid van Plasmocin™ op diverse zoogdiercellijnen die zijn geïnfecteerd met mollicutes, vergeleken met veelgebruikte antibiotica in celcultuur: een lokale ervaring. Cytotechnology. 2011;63:609-20
[21] Kragh Hansen U. Moleculaire aspecten van ligandbinding aan serumalbumine. Pharmacol Rev. 1981;33:17-53