
HepG2-solut – maksasyövän tutkimuksen resurssi
Hep-G2 on ihmisen maksasyöpäsolulinja, joka on peräisin 15-vuotiaan valkoihoisen miehen maksakudoksesta, jolla oli maksasyöpä. Näitä soluja käytetään usein lääkeaineenvaihduntaa ja maksatoksisuutta koskevissa tutkimuksissa. Vaikka HepG2-soluilla on korkea proliferaatioaste ja epiteelin kaltainen ulkonäkö, ne eivät ole tuumorigeenisiä ja suorittavat erilaisia erilaistuneita maksan toimintoja. Vuonna 1975 tutkijat eristivät HepG2-solut hepatosellulaarisesta karsinoomasta, jolloin siitä tuli ensimmäinen maksasolulinja, jolla oli hepatosyyttien kriittiset ominaisuudet. Toisin kuin aiemmin perustettu SK-Hep1-solulinja, josta puuttuvat olennaiset maksasolujen merkkiaineet, HepG2-solut voivat erittää erilaisia plasmaproteiineja ja tarjoavat arvokkaan mallin ihmisen hepatosyyttien solupinnan alueiden solunsisäisen dynamiikan tutkimiseen. Nämä solut ovat epiteelin kaltaisia, niiden kromosomiluku on tyypillisesti 55, ja niitä voidaan stimuloida ihmisen kasvuhormonilla.
- Kasvatusväliaine
- Katso tuotesivu
- Kaksinkertaistumisaika
- Katso tuotesivu
- Kasvutyyppi
- Adherentti
- Bioturvallisuustaso
- BSL-1
- Saatavana
- Cytion — Tilaa HepG2
HepG2-solujen ominaisuudet
Ensisijaisten hepatosyyttien tyypillinen muoto on kuutiomainen, ja niissä on yleensä kaksi tumaa. Sen sijaan HepG2-soluilla on epiteelin kaltainen morfologia, yksi tuma ja kromosomimäärä, joka vaihtelee solua kohti 48:sta 54:ään. Vaikka HepG2-solut voivat muodostaa jopa 25 % solun kokonaisproteiinista, niiden koko on normaaleja hepatosyyttejä suurempi, ja ne muodostavat noin 10 % solun kokonaisproteiinista. Soluproteiinit ovat solun sisällä keskeisiä toimijoita, jotka suorittavat geenien määrittelemiä toimintoja.
Kasvainsolut, mukaan lukien ne, joilla on epänormaali kromosomimäärä, osoittavat usein ytimien määrän kasvua, jopa seitsemään solua kohti. Korkean in vitro -erilaistumisasteensa ansiosta HepG2-solut tarjoavat ihanteellisen mallin ihmisen hepatosyyttien sappikanavien, sinusoidien kalvoproteiinien ja lipidien solunsisäisen kuljetuksen ja dynamiikan tutkimiseen.
HepG2-solun keskimääräinen halkaisija on noin 10–20 µm, mikä on pienempi kuin 15 µm:n halkaisijaltaan olevan hepatosyytin, mutta samanlainen kuin hepatoblastooma (HB) -kasvainsolujen, joiden halkaisija on 10–20 µm.
HepG2-genetiikka
Hep-G2-solulinjassa esiintyy useita translokaatioita, mukaan lukien kromosomien 1 ja 21 lyhyiden käsivarsien väliset translokaatiot, kromosomien 2, 16 ja 17 trisomiat sekä kromosomin 20 tetrasomia. Myös kromosomin 4q3-alueen menetys on havaittu, ja se liittyy translokaatioon t(1;4), jota esiintyy usein hepatoblastoomassa (HB), sekä muihin kromosomipoikkeavuuksiin, kuten trisomioihin 2 ja 20. HepG2-solujen kromosomien lukumäärä vaihtelee 50:stä 60:een, mikä viittaa hyperdiploidiin karyotyyppiin, kun taas joissakin tapauksissa kromosomien lukumäärä on yli 100 ja niille on ominaista tetraploidi laajentuminen. HepG2-solut sisältävät noin 7,5 pg DNA:ta, mikä on 15 % enemmän kuin keskimääräisessä somaattisessa solussa. Vertailun vuoksi primaariset hepatosyytit ovat kuutiomaisia ja sisältävät tyypillisesti kaksi tumaa [1].
HepG2-solujen mutaatioprofiili
HepG2-solulinjassa on TERT-promoottorialueen mutaatio C228T, jota esiintyy myös hepatosellulaarisessa karsinoomassa (HCC) ja hepatoblastoomassa (HB). Tämä mutaatio edistää ikuistumista suojaamalla telomeerejä syöpäsoluissa. Lisäksi HepG2-soluissa esiintyy villityyppistä TP53-geeniä, joka on kriittinen geeni ihmisen syövän tukahduttamisessa, sillä se vaikuttaa solusyklin pysähtymiseen, apoptoosiin ja ikääntymiseen. Tämän geenin mutaatiot voivat edistää solujen lisääntymistä.
HepG2-solut osallistuvat useisiin reitteihin, mukaan lukien solujen kasvun säätelyn häiriöt, selviytymisreitit, kuten sikiön ja alkion HB, sekä Wnt/β-kateniini-reitti. Lisäksi solulinjalla on tyypillinen CTNNB1-geenin kolmannen eksonin deleetio, joka on identtinen epiteelityyppisessä HB:ssä havaittavan kanssa [2,3].

Katsaus HepG2-maksasyöpäsoluihin maksatutkimuksessa
Ihmisen hepatomasta peräisin olevat HepG2-solut ovat tulleet korvaamattomaksi työkaluksi maksan toimintojen ja sairauksien, mukaan lukien hepatosellulaarisen karsinooman, tutkimuksessa. Nämä maksasolulinjat antavat tietoa ihmisen makrosolujen soluvasteista erilaisissa kokeellisissa olosuhteissa. Lusiferaasi-raportointiplasmidien käyttö HepG2-soluissa on ollut erityisen tehokasta geenien ilmentymisen ja solutransfektioiden seurannassa, jotka ovat keskeisiä aineenvaihdunnan tutkimuksessa, kuten etanolin vaikutusten tutkimuksessa maksasoluihin.
Virusinfektioiden ja maksasairauksien tutkimus HepG2-soluilla
HepG2:n ja Huh7:n kaltaiset ikuistetut maksasyöpäsolulinjat ovat välttämättömiä virusinfektioiden tutkimuksessa, sillä ne osoittavat hepatiitti D:n (HDV) täydellisen solusyklin replikaation ja hepatiitti B:n (HBV) ilmentymisen [5,6]. Samalla HepaRG-solulinjat ovat keskeisessä roolissa HBV:n soluunpääsymekanismien selvittämisessä [7]. HepG2-soluja käytetään myös erilaisten ihmisen maksasairauksien tutkimiseen, progressiivisesta familiaalisesta intrahepaattisesta kolestaasista (PFIC) ja Dubin-Johnsonin oireyhtymästä sytotoksisiin ja genotoksisiin aineisiin liittyviin ympäristö- ja ruokavaliotutkimuksiin sekä lääkkeiden kohdentamiseen ja maksasyövän syntyyn liittyvään tutkimukseen [8,9]. Niiden käyttö ulottuu bio-keinotekoisten maksa-laitteiden kokeisiin.
HepG2-solujen vuorovaikutus biomateriaalien kanssa kudostekniikassa
HepG2-solujen vuorovaikutus erilaisten biomateriaalien kanssa on keskeistä kudostekniikassa. Kolloidisen koettimen tekniikan kaltaiset menetelmät auttavat ymmärtämään näitä vuorovaikutuksia mittaamalla solujen adheesio-ominaisuuksia, jotka ovat elintärkeitä solujen elinkelpoisuuden määrittämisessä tukirakenteiden ja tarkkojen maksakudosmallien kehittämiseksi.
Solujen käyttäytyminen ja innovaatiot HepG2-pohjaisissa malleissa
HepG2-pohjaisten mallien solukäyttäytymisen tutkiminen on ratkaisevan tärkeää maksasairauksien tutkimuksessa. Kolmiulotteisten sfääristen soluviljelmien kehitys on johtanut HepG2-solusfäärien luomiseen, jotka tarjoavat fysiologisesti merkityksellisemmän mallin, joka heijastaa tarkasti normaaleja hepatosyyttejä. Nämä 3D-mallit, joiden aineenvaihdunta on vilkkaampaa, viittaavat HepG2-solujen potentiaaliin toimia hepatoblastooman mallina, ja ne ovat tärkeitä syövän hoitotutkimuksessa, erityisesti maksakasvainten simuloinnissa ja uusien hoitomenetelmien testaamisessa [10-12].
HepG2:n vertailu ja ominaisuudet muihin kasvainsolulinjoihin nähden
HepG2 on yksi laajimmin käytetyistä maksakasvainsolulinjoista, joka on valittu sen laajan soveltuvuuden vuoksi tieteelliseen tutkimukseen noin 40 saatavilla olevan maksakasvainsolulinjan joukosta [13]. Huolimatta siitä, että tiettyjen sytokromi P450 -entsyymien ilmentyminen on heikkoa tai puuttuu kokonaan verrattuna normaaleihin hepatosyytteihin, HepG2:n metabolinen profiili on edistänyt pyrkimyksiä muokata solulinjaa parempien lääkeaineen metaboliaa koskevien tutkimusten tekemiseksi [13]. Verrattuna syöpäsolulinjoihin, kuten MCF7, PC3, 143B ja HEK293, HepG2-soluilla on ainutlaatuiset aminohappopitoisuusprofiilit, jotka vaikuttavat merkittävästi proteiinisynteesiin ja -eritykseen, mikä korostaa niiden ainutlaatuisia aineenvaihduntareittejä [14].
Maksasairauksien tutkimusta HepG2-solulinjan avulla
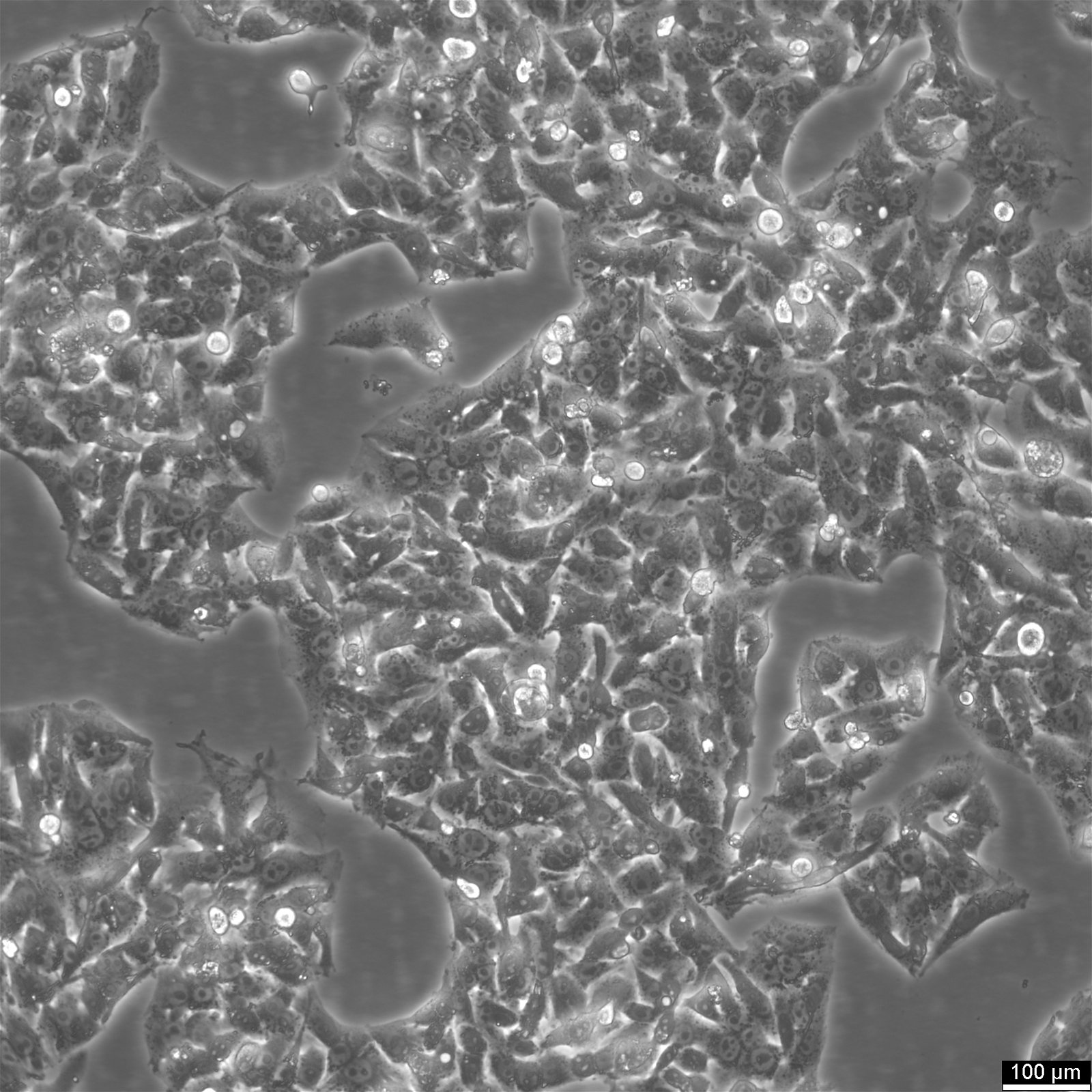
HepG2-solujen siirrostaminen
Seuraavassa on viisi vaihetta kiinnittyneiden solujen poistamiseksi soluviljelypulloista Accutase-entsyymillä:
- Poista elatusaine soluviljelypullosta ja huuhtele kiinnittyneet solut kalsiumia ja magnesiumia sisältämättömällä PBS-liuoksella. Käytä 3–5 ml PBS-liuosta T25-pulloihin ja 5–10 ml T75-pulloihin.
- Lisää Accutase soluviljelypulloon, 1–2 ml T25-pulloa kohti ja 2,5 ml T75-pulloa kohti. Varmista, että Accutase peittää koko solukerroksen.
- Inkuboi pulloa huoneenlämmössä 8–10 minuuttia.
- Suspendoi solut varovasti uudelleen käyttämällä 10 ml tuoretta elatusainetta.
- Sentrifugoi suspendoidut solut 5 minuuttia 300 x g:n nopeudella, suspendoi ne uudelleen tuoreeseen elatusaineeseen ja siirrä ne uusiin pulloihin, jotka sisältävät tuoretta elatusainetta.
HepG2-solujen tulevaisuudennäkymät
Pyrkimys HepG2-solulinjan täyden potentiaalin hyödyntämiseen jatkuu, ja sytokromien ilmentymisen lisäämisessä on saavutettu uraauurtavaa edistystä. Tutkijat tutkivat myös kolmiulotteisten sferoidisten soluviljelmien mahdollisuuksia, jotka tarjoavat fysiologisesti merkityksellisemmän järjestelmän. Metabolinen aktiivisuus, mukaan lukien sytokromit, on huomattavasti korkeampi kolmiulotteisissa sfäärisissä HepG2-malleissa kuin kaksiulotteisissa soluissa, mikä vie meitä lähemmäksi normaalien hepatosyyttien kaltaisen mallin luomista. Lisäksi solupintaproteiinien virheellisen jakautumisen taustalla olevien dynaamisten prosessien tutkiminen voi avata tien maksa-sairauksien parempaan ymmärtämiseen.
HepG2-solut: niiden roolin ja erityispiirteiden ymmärtäminen biolääketieteellisessä tutkimuksessa – Usein kysyttyjä kysymyksiä
Lähteet
- Vyas, R.C., Darroudi, F., Natarajan, A.T. Säteilyn aiheuttamat kromosomien katkeamiset ja uudelleenliittymiset ihmisen lymfosyyttien interfasi-metafaasikromosomeissa, Mutat Res, 1991; 249(1):29-35.
- Woodfield, S.E., Shi, Y., Patel, R.H., Chen, Z., Shah, A.P., Srivastava, R.K., Whitlock, R.S., Ibarra, A.M., Larson, S.R., Sarabia, S.F., et al. MDM4:n estäminen: uusi terapeuttinen strategia P53:n aktivoimiseksi hepatoblastoomassa. Sci. Rep. 2021, 11, 2967.
- Hussain, S.P., Schwank, J., Staib, F., Wang, X.W., Harris, C.C. TP53-mutaatioita ja hepatosellulaarinen karsinooma: näkemyksiä maksasyövän etiologiasta ja patogeneesistä. Oncogene 2004.
- Schicht, G., Seidemann, L., Haensel, R., Seehofer, D., Damm, G. Kriittinen tutkimus hepatomisolulinjojen HepG2 ja Huh7 käyttökelpoisuudesta malleina resektoitavan hepatosellulaarisen karsinooman metaboliselle esittämiselle. Cancers 2022, 14(17), 4227.
- Verrier, E.R., Colpitts, C.C., Schuster, C., Zeisel, M.B., Baumert, T.F. Soluviljelymallit B- ja D-hepatiittivirustartuntojen tutkimiseen. Viruses 2016, 8, 261.
- Verrier, E.R., Colpitts, C.C., Bach, C., Heydmann, L., Weiss, A., Renaud, M., Durand, S.C., Habersetzer, F., Durantel, D., AbouJaoudé, G., et al. Kohdennettu toiminnallinen RNA-interferenssiseulonta paljastaa glypikaani 5:n hepatiitti B- ja D-virusten tunkeutumistekijäksi. Hepatology 2016, 63, 35–48.
- Gripon, P., Rumin, S., Urban, S., Le Seyec, J., Glaise, D., Cannie, I., Guyomard, C., Lucas, J., Trepo, C., Guguen-Guillouzo, C. Hepatiitti B -viruksen aiheuttama infektio ihmisen hepatomasolulinjassa. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660.
- Mersch-Sundermann, V., Knasmüller, S., Wu, X.J., Darroudi, F., Kassie, F. Ihmisperäisen maksasolulinjan käyttö soluja suojaavien, antigenotoksisten ja kogenotoksisten aineiden havaitsemisessa. Toxicology. 2004; 198(1–3): 329–340.
- Fanelli, A. HepG2 (maksan hepatosellulaarinen karsinooma): soluviljely. HepG2. Haettu 3. joulukuuta 2017.
- Xuan, J., Chen, S., Ning, B., Tolleson, W.H., Guo, L. Sytokromi P450:iä ilmentävien HepG2-peräisten solujen kehittäminen aineenvaihduntaan liittyvän lääkeaineiden aiheuttaman maksatoksisuuden arvioimiseksi. Physiol. Behav. 2017, 176, 139–148.
- Ooka, M., Lynch, C., Xia, M. In vitro -metabolian aktivoinnin soveltaminen suurikapasiteettisessa seulonnassa. Int. J. Mol. Sci. 2020, 21, 8182.
- Huang, L., Coughtrie, M.W.H., Hsu, H. Dehydroepiandrosteronisulfotransferaasigeenin alisäätely ihmisen maksasyövässä. Mol. Cell. Endocrinol.
- Zhu, Z., Hao, X., Yan, M., et al. Syövän kantasolut ja esisolut ovat erittäin runsaasti edustettuina CD133 + CD44 + -populaatiossa maksasyövässä. Int J Cancer. 2010; 126:2067-2078.
- Arbus, C., Benyamina, A., Llorca, P.-M., Baylé, F., Bromet, N., Massiere, F., Garay, R.P., Hameg, A. Syamematsiinin metaboliaan osallistuvien ihmisen sytokromi P450 -entsyymien karakterisointi. Eur J Pharm Sci. 2007 Dec;32(4-5):357-66.