
, L-alanil-L-glutamin oldat")
Térfogat: 100 ml
Tárolás: ≤ –15 °C
Sterilitás: Steril szűrt
A stabil glutaminoldat (L-alanil-L-glutamin, 200 mM) egy rendkívül stabil dipeptid formájú L-glutamin, amelyet a sejtkultúra-táptalajokban a hagyományos L-glutamin közvetlen helyettesítésére fejlesztettek ki. Az L-glutamin egy esszenciális aminosav és a tenyésztett sejtek fő energiaforrása, amely kritikus szerepet játszik a sejtek növekedésében, anyagcseréjében és fehérjeszintézisében.
Alkalmazás és előnyök
A standard folyékony táptalajokban az L-glutamin 37 °C-on viszonylag gyorsan lebomlik, ami toxikus melléktermékek, például ammóniumionok képződéséhez vezet, amelyek negatívan befolyásolhatják a sejtek életképességét és a kísérleti eredményeket. A stabil glutamin leküzdíti ezt a korlátozást azzal, hogy nem bomló dipeptid formát biztosít, amely tenyésztési körülmények között is sértetlen marad.
A sejtek enzimatikusan hasítják a dipeptidkötést, hogy szükség szerint felszabadítsák az L-glutamint, biztosítva ezzel a folyamatos friss utánpótlást, miközben megakadályozzák a káros hulladéktermékek felhalmozódását. Ez különösen előnyös a hosszú távú sejtkultúrák és a nagy sűrűségű növekedési rendszerek esetében.
Összetétel és felhasználás
Az L-alanil-L-glutamin oldatot sejtkultúra-minőségű vízben készítik elő 200 mM koncentrációban, és szennyeződésre érzékeny alkalmazásokhoz steril szűrővel szűrik. A kísérleti követelményeknek megfelelően közvetlenül teljes táptalajba hígítható. A termék stabilitásának megőrzése érdekében ≤ –15 °C-on tárolandó, és kerülni kell az ismételt fagyasztás-olvasztás ciklusokat.
Kizárólag kutatási célra. Diagnosztikai vagy terápiás eljárásokhoz nem használható. Emberben vagy állatokban nem alkalmazható.
")
Térfogat: 100 ml
Tárolás: +2 °C és +8 °C között
Sterilitás: Steril szűrt
A HEPES pufferoldat (1 M), más néven N-2-hidroxi-etil-piperazin-N-2-etán-szulfonsav, egy zwitterionos szerves pufferközeg, amelyet széles körben használnak sejtkultúra-táptalajokban. Úgy tervezték, hogy stabil pH-értéket biztosítson a 6,7 és 8,6 közötti fiziológiai tartományban, támogatva az optimális sejtfunkciót in vitro alkalmazások során.
Alkalmazás és előnyök
A HEPES megbízható pufferkapacitást biztosít a sejtkultúra-rendszerekben, különösen akkor, ha a sejteket CO₂-inkubátoron kívül kezelik. 10–25 mM HEPES hozzáadása a tenyésztőközeghez fokozott pH-stabilitást biztosít hosszabb manipulációs időszakok alatt, segítve az állandó kísérleti feltételek fenntartását.
Ez a puffer membránáteresztő, minimálisan zavarja a biokémiai reakciókat, és erős kémiai és enzimatikus stabilitást mutat. Ezek a tulajdonságok alkalmassá teszik széles körű sejtkultúra
- és biokémiai alkalmazásokra.
Összetétel és felhasználás
A megoldás 1 M koncentrátum formájában kerül forgalomba, sejtkultúra-minőségű vízben elkészítve, és steril szűrésen esik át, hogy szennyeződésre érzékeny környezetben is használható legyen. Az alkalmazás követelményeinek megfelelően a kívánt munkakoncentrációra hígítható. +2 °C és +8 °C között tárolandó, és a termék integritásának megőrzése érdekében aszeptikus körülmények között kell kezelni.
Kizárólag kutatási célra. Diagnosztikai vagy terápiás eljárásokban nem használható. Emberben vagy állatokban nem alkalmazható.
, 250 g/l")
Térfogat: 50 ml
Tárolás: +2 °C és +8 °C között
Sterilitás: Steril szűrt
A D-(+)-glükóz oldat (dextróz oldat) egy steril, használatra kész kiegészítő, amely a sejtek anyagcseréjének központi eleme, a természetesen előforduló D-(+)-glükóz cukrot tartalmazza. A glükóz olyan alapvető biológiai folyamatokban vesz részt, mint az energia termelés, a glikoziláció és a sejtek szerkezetéhez és működéséhez hozzájáruló glikánok képződése.
Alkalmazás és előnyök
Ez a glükózoldat széles körben használatos kiegészítőként sejtkultúra-táptalajokban és számos sejt
- és molekuláris biológiai alkalmazásban. Elsődleges szén
- és energiaforrásként a glükóz támogatja a sejtek növekedését, szaporodását és anyagcsere-aktivitását. A bioszintézis útjaiban betöltött szerepe miatt kritikus fontosságú a normális sejtfiziológia és a kísérleti konzisztencia fenntartásában.
Összetétel és felhasználás
Az oldat 250 g/l glükóz magas koncentrációban kerül forgalomba, ami lehetővé teszi a tenyésztőközegbe való rugalmas hígítást a kísérleti követelményeknek megfelelően. Steril szűrésen esik át, hogy szennyeződésre érzékeny alkalmazásokhoz is alkalmas legyen. +2 °C és +8 °C között tárolandó, és a termék minőségének és stabilitásának megőrzése érdekében aszeptikusan kell kezelni.
Kizárólag kutatási célra. Diagnosztikai vagy terápiás eljárásokhoz nem használható. Emberben vagy állatokban nem alkalmazható.
, 100-szoros oldat")
Térfogat: 10 ml
Tárolás: +2 °C és +8 °C között
Sterilitás: Szteril szűrt
Az inzulin-transferrin-szelén (ITS) oldat (100x) egy kémiailag meghatározott kiegészítő, amelyet széles körű sejtkultúra-alkalmazásokhoz fejlesztettek ki. Leggyakrabban alapsejtkultúra-táptalajok adalékaként használják, hogy támogassa a sejtek növekedését csökkentett szérumtartalmú vagy szérummentes körülmények között.
Alkalmazás és előnyök
ITS kiegészítőnk biztosítja a szérummentes táptalajok optimális működéséhez szükséges alapvető összetevőket. A hagyományos tápközeg ITS-szel történő kiegészítésével jelentősen csökkenthető a számos sejtvonal rutin fenntartásához szükséges borjúszérum (FBS) mennyisége. Ez segít minimalizálni a szérumhasználattal járó variabilitást, miközben biztosítja a sejtek egyenletes növekedését és életképességét.
Az inzulin támogatja a kulcsfontosságú tápanyagok sejtbejutását és anyagcseréjét, a transzferrin elősegíti a vas szállítását, a szelén pedig hozzájárul az antioxidáns védelemhez és az enzimaktivitáshoz. Ezek a komponensek együttesen elősegítik a kiegyensúlyozott sejtanyagcserét és javítják a reprodukálhatóságot meghatározott tenyésztési rendszerekben.
Összetétel és felhasználás
Az inzulin-transferrin-szelén (ITS) 100-szoros koncentrátumként, fenolvörös nélküli Earle's Balanced Salt Solution (EBSS) oldatban kerül forgalomba. Szabványos alkalmazásokhoz 1:100 arányban hígítsa a megfelelő alapközegbe az ajánlott munkakoncentráció elérése érdekében. A termék stabilitásának és sterilitásának megőrzése érdekében +2 °C és +8 °C között tárolja, és aszeptikus körülmények között kezelje.
Kizárólag kutatási célra. Diagnosztikai vagy terápiás eljárásokhoz nem alkalmazható. Emberre vagy állatra nem alkalmazható.
")
Térfogat: 5 ml
Tárolás: +2 °C és +8 °C között
Sterilitás: Szteril szűrt
A humán rekombináns inzulinoldat egy kémiailag meghatározott kiegészítő, amelyet általában emlős sejtvonalak, többek között kínai hörcsög petefészek (CHO) sejtek tenyésztéséhez használnak. Ez a sejtkultúra-minőségű oldat Saccharomyces cerevisiae-ben expresszált rekombináns humán inzulint tartalmaz, ami biztosítja a magas tisztaságot és az állandó teljesítményt a kutatási alkalmazásokban.
Alkalmazás és előnyök
Az inzulint rutinszerűen adják hozzá a szérummentes és kémiailag definiált táptalajokhoz a sejtek növekedésének és termelékenységének elősegítése érdekében. Kulcsfontosságú szabályozó hormonként az inzulin támogatja a glükóz, az aminosavak és a zsírsavak sejtek általi felvételét, hasznosítását és tárolását. Gátolja továbbá a glikogén, a fehérjék és a lipidek lebontását, hozzájárulva ezzel a sejtek életképességének javulásához és az anyagcsere stabilitásához a tenyésztési rendszerekben. A kémiailag definiált készítmény elősegíti a reprodukálhatóságot és minimalizálja a variabilitást az érzékeny sejtkultúra-munkafolyamatokban.
Biológiai tulajdonságok és felhasználás
Az inzulin egy kétláncú polipeptid hormon, amelyet a hasnyálmirigy szigeteinek β-sejtjei termelnek természetes úton. Molekulatömege körülbelül 5800 Da. Az α
- és β-láncokat két láncok közötti diszulfidkötés köti össze, az α-lánc pedig egy láncon belüli diszulfidkötést tartalmaz. Sejtkultúra-alkalmazások esetén az oldatot aszeptikus körülmények között kell kezelni, és +2 °C és +8 °C között kell tárolni a stabilitás és a teljesítmény fenntartása érdekében.
Kizárólag kutatási célra. Diagnosztikai vagy terápiás eljárásokhoz nem használható. Emberben vagy állatokban nem használható.
")
Térfogat: 100 ml
Tárolás: +2 °C és +8 °C között
Sterilitás: Steril szűrt
A nátrium-piruvát oldat (100 mM) egy steril, használatra kész kiegészítő, amelynek célja, hogy további, könnyen hozzáférhető energiaforrást biztosítson a sejtkultúra táptalajához. A nátrium-piruvát kulcsszerepet játszik a sejtek energia-anyagcseréjében, és támogatja a metabolikusan aktív és gyorsan szaporodó sejtek, például a tumorsejtek növekedését. A kiegészítés javíthatja a sejtek életképességét és segíthet fenntartani az anyagcsere stabilitását a tenyésztési rendszerekben.
Alkalmazás és előnyök
Ezt az oldatot széles körben használják a rutin sejtkultúrában a táptalaj piruváttal való dúsítására és az optimális növekedési feltételek elősegítésére. Támogatja az ATP-termelést, segíthet az oxidatív stressz csökkentésében, és hozzájárul a tenyésztett sejtek anyagcsere-teljesítményének javításához. A sejtkultúra-minőségű vízben gyártott és steril szűrt termék biztosítja a kutatási munkafolyamatok állandó minőségét és reprodukálhatóságát.
Használat és kompatibilitás
A legtöbb sejtkultúra-alkalmazáshoz ajánlott végső koncentráció 1 mM nátrium-piruvát, amelyet a 100 mM-es törzsoldat 1:100 arányú hígításával lehet elérni a teljes tenyésztőközegben. Az oldat számos alapközeggel és emlős sejtvonalakkal kompatibilis. A termék stabilitásának megőrzése érdekében +2 °C és +8 °C között tárolandó, és szennyeződéstől védve.
Kizárólag kutatási célra. Diagnosztikai vagy terápiás eljárásokhoz nem használható. Emberben vagy állatokban nem alkalmazható.
")
Térfogat: 100 ml Tárolás: ≤-15°C Sterilitás: Steril szűrt
Az Antibiotikum/Antimikotikum oldat (100x) egy steril, felhasználásra kész koncentrátum, amelyet úgy terveztek, hogy csökkentse a mikrobiális szennyeződés kockázatát a sejtkultúrában és a kapcsolódó laboratóriumi alkalmazásokban. Ez a 100x oldat a penicillin, a streptomicin és az amfotericin B jól bevált kombinációját tartalmazza, amely széles spektrumú antimikrobiális aktivitást biztosít Gram-pozitív és Gram-negatív baktériumok, élesztők és fonalas gombák ellen. A készítmény alkalmas eukarióta sejttenyészetekben, bakteriális táptalajokon és más, szennyeződésre érzékeny rendszerekben történő felhasználásra, támogatva a tiszta és következetes laboratóriumi műveleteket.
Alkalmazás és előnyök A rutin kutatási protokollokhoz optimalizált oldatot széles körben használják a sejtkultúra munkafolyamatok aszeptikus körülményeinek fenntartására. Megbízható teljesítményt nyújt a szennyeződésre érzékeny környezetekben, segítve a kutatókat a mikrobák elszaporodásának kockázatának csökkentésében anélkül, hogy veszélyeztetné a sejtek egészségét vagy a kísérleti reprodukálhatóságot. A sterilszűrt formuláció kiküszöböli a további szolubilizációs lépések szükségességét, támogatva a racionalizált médiumkészítést és csökkentve a napi laboratóriumi eljárások variabilitását.
Használat és kompatibilitás A standard munkakoncentrációk eléréséhez hígítsa az oldatot 1:100 arányban a teljes táptalajhoz. A termék kompatibilis az emlős sejtvonalak és alapközegek széles skálájával. A folyamatos készletelérhetőségnek köszönhetően a kutatók számára előnyös a megbízható ellátás folyamatossága és az egyszerűsített logisztikai tervezés. Az oldatot ≤ -15 °C-on kell tárolni, és a stabilitás megőrzése érdekében védeni kell az ismételt fagyasztási-olvasztási ciklusoktól. Kizárólag kutatási célú felhasználásra. Nem használható diagnosztikai vagy terápiás eljárásokban. Nem használható embereken vagy állatokon.
")
Térfogat: 100 ml Tárolás: sterilitás: +2°C és +8°C között: Steril szűrt
AMEM nem esszenciális aminosavak (100x) egy steril kiegészítés, amelyet a sejtnövekedés és életképesség fokozására terveztek emlős sejttenyésztő rendszerekben. A készítmény a standard Minimum Essential Mediumban (MEM) található nem esszenciális aminosavak 100x-os koncentrátumának felel meg, lehetővé téve az alapközegek közvetlen kiegészítését minimális előkészítéssel.
Alkalmazás és előnyök Ez a kiegészítés kiegészítő aminosavkészletet biztosít a gyorsan proliferáló sejtek vagy olyan sejtvonalak számára, amelyek elvesztették a nem esszenciális aminosavak de novo szintézisének képességét. A bioszintézis metabolikus terhének enyhítésével támogatja a jobb növekedési kinetikát, a hosszabb életképességet és a nagyobb kísérleti konzisztenciát
- különösen a tápanyagérzékeny vagy nagy sűrűségű kultúrákban.
Összetétel és felhasználás Az oldat glicint, L-alanint, L-aszparagint, L-aszparaginsavat, L-glutaminsavat, L-prolint és L-szerint tartalmaz. Kompatibilis a MEM-mel és a legtöbb más standard táptalajjal. Használathoz hígítsa 1:100 arányban a végleges táptalajba. Ez a termék sterilszűrt és további kezelési lépések nélkül felhasználásra kész. Kizárólag kutatási célú felhasználásra. Nem használható diagnosztikai vagy terápiás eljárásokban. Nem használható embereken vagy állatokon.

Az Accutase egy használatra kész, steril szűrt sejtleválasztó oldat, amelyet a tripszin/EDTA kíméletes alternatívájaként fejlesztettek ki a tapadó emlős sejtek standard szövetkultúra-műanyag edényekről és tapadásgátló bevonattal ellátott felületekről történő leválasztására. Proteolitikus és kollagenolitikus enzimaktivitást ötvöz egy kiegyensúlyozott sóoldatban, hogy hatékony, ugyanakkor kontrollált leválasztást biztosítson, megőrizve a sejtfelszíni fehérjéket, és támogatva a magas posztpasszázs-életképességet és a gyors újratapadást.
Az Accutase összetétele Dulbecco foszfátpufferolt sóoldatán (DPBS) alapul, EDTA-val és fenolvörössel, mint vizuális pH-indikátorral. Az enzimek nem emlős és nem bakteriális eredetűek, ezért az Accutase különösen alkalmas őssejtkutatásra, vakcina-előállítási folyamatokra és minden olyan alkalmazásra, ahol az állati vagy mikrobiális eredetű szennyeződéseket minimálisra kell csökkenteni. Az oldat 37 °C-on öninhibálódik, ezért a leválasztás után nincs szükség semlegesítő reagensre vagy szérumtartalmú táptalajra – a sejteket közvetlenül friss táptalajba lehet átvinni.
Főbb jellemzők
Használatra kész, 1x-es, steril szűrt folyadék – hígításra vagy rekonstitúcióra nincs szükség
Kombinált proteolitikus és kollagenolitikus enzimaktivitás a kíméletes disszociáció érdekében
Minden tétel standardizált, meghatározott diszociációs aktivitással a tételek közötti konzisztencia érdekében
Nem emlős és nem bakteriális enzim eredetű
37 °C-on öninhibálódik – nincs szükség semlegesítő oldatra
Dulbecco PBS-ben EDTA-val készítve
Fenolvörös vizuális pH-indikátorként
pH 6,8 – 7,8
Jellemző alkalmazások
Az Accutase kíméletesen disszociálja a legkülönbözőbb adhezív és érzékeny sejttípusokat, beleértve az emberi embrionális őssejteket (hESCs), az emberi indukált pluripotens őssejteket (iPSCs), az idegsejteket, az elsődleges idegsejteket, valamint a rutinszerűen tenyésztett adhezív sejtvonalakat, mint például a HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 és A549. Tipikus felhasználási területek:
Adhéziós emlős sejtek rutinszerű szubkultiválása és átültetése
hESC-k, iPSC-k és más érzékeny vonalak kíméletes egysejtes disszociációja
Minták előkészítése áramlási citometriához és FACS-elemzéshez
Olyan sejtfelszíni markerek elemzése, ahol az epitóp integritása fontos
Sejtvándorlás, proliferáció és apoptózis vizsgálatok
Nyugalmi állapot vizsgálata szérumhiány és onkogén transzfekciós vizsgálatok segítségével
Tumorsejtek és idegcsíra-sejtek migrációs vizsgálatai
Termelés felfuttatása bioreaktoros munkafolyamatokban
Rutin munkákhoz körülbelül 10 ml Accutase-t alkalmazzon 75 cm2 tenyésztési felületre, és inkubálja 5–10 percig szobahőmérsékleten. Az optimális inkubációs időt minden sejtvonal esetében meg kell határozni, és az nem haladhatja meg az egy órát. Az adagolás előtt öblítse le a sejtréteget Ca2+/Mg2+-mentes sóoldattal, például kalcium
- és magnéziummentes DPBS-sel, hogy eltávolítsa a maradék szérumot és a kétértékű kationokat.
Kezelés és tárolás
A bontatlan palackot -15 °C-on vagy annál alacsonyabb hőmérsékleten fagyassza le. Szobahőmérsékleten vagy egy éjszakán át +2 °C és +8 °C között olvassza fel. Ne olvassza fel az Accutase-t 37 °C-os vízfürdőben, mivel a magas hőmérséklet csökkenti az enzimaktivitást. Felolvasztás után az oldat +2 °C és +8 °C között legfeljebb 2 hónapig tárolható; ne tárolja szobahőmérsékleten. Az alkalmazás előtt ne melegítse elő a reagenset 37 °C-ra – adja hozzá közvetlenül a mosott sejtekhez szobahőmérsékleten. A hosszú távú tárolhatóság érdekében az ismételt felolvasztási ciklusok elkerülése érdekében egyszer használatos adagolás ajánlott. Mindig aszeptikus körülmények között dolgozzon.
Minőség
Szigorú minőségi előírások szerint gyártva. Az Accutase minden tételét szteril szűrőn szűrik, és szterilitás, pH, megjelenés és disszociációs aktivitás szempontjából tesztelik, hogy tételenkénti konzisztens, reprodukálható teljesítményt biztosítsanak.
Termékjellemzők
Specifikáció
Részlet
TerméktípusSejtleválasztó / disszociációs reagens
FormátumSteril szűrt folyadék, használatra kész
Térfogat100 ml
Munkakoncentráció1x (használatra kész)
EnzimaktivitásKombinált proteolitikus és kollagenolitikus
Enzim eredeteNem emlős és nem bakteriális
Puffer rendszerDulbecco PBS EDTA-val
pH-indikátorFenolvörös
pH-tartomány6,8 – 7,8
KinézetTiszta, halványvörös vagy narancssárga oldat
Tárolási hőmérséklet-15 °C vagy annál alacsonyabb
Stabilitás felolvasztás utánLegfeljebb 2 hónap +2 °C és +8 °C között
Ajánlott felhasználási mennyiség~10 ml 75 cm2 tenyésztési felületre
Jellemző inkubációs idő5–10 perc szobahőmérsékleten
Szállítási feltételekSzárazjégen fagyasztva
RendeltetésKizárólag kutatási célra és további gyártásra
Összetétel (literenkénti összetétel)
Összetevő
Koncentráció (mg/l)
Szervetlen sók
Nátrium-klorid (NaCl)8000,00
Dinátrium-hidrogén-foszfát (Na2HPO4)1150,00
Kálium-klorid (KCl)200,00
Kálium-dihidrogén-foszfát (KH2PO4)200,00
Egyéb összetevők
EDTA · 4Na (tetranátrium-EDTA)220,00
Fenolvörös3,00
Saját fejlesztésű enzimkeverék (proteolitikus és kollagenolitikus hatással)1x
Az Accutase az Innovative Cell Technologies, Inc. bejegyzett védjegye.
, w: 2 mM L-Glutamin, w: 2,2 g/L NaHCO3, w: EBSS")
Ez a használatra kész, steril szűrt folyékony készítmény Earle’s Balanced Salt Solution (EBSS) oldattal, 2 mM L-glutaminnal, D-glükózzal (1,0 g/l) és 2,2 g/l nátrium-hidrogén-karbonáttal (NaHCO3) van kiegészítve, így alkalmas CO2-szabályozott inkubátorban (jellemzően 5 % CO2) történő használatra. A készítményben található fenolvörös pH-indikátorként működik, lehetővé téve a tápközeg állapotának kényelmes vizuális ellenőrzését a sejtkultúra során.
Főbb jellemzők
Klasszikus Eagle’s MEM készítmény Earle’s Balanced Salt Solution (EBSS)
2 mM L-glutamin tartalommal – azonnal felhasználható
2,2 g/l nátrium-hidrogén-karbonát – 5 %-os CO2-inkubációra pufferelve
D-glükóz (1,0 g/l) mint elsődleges szénforrás
Fenolvörös pH-indikátorral
HEPES és nátrium-piruvát nélkül
Steril szűrt folyékony táptalaj, használatra kész
pH 7,0 – 7,6
Jellemző alkalmazások
Az EMEM számos emlős sejtvonal tenyésztését támogatja, beleértve a HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 és számos primer sejtet. Gyakori alkalmazások:
Adherens sejtvonalak rutin karbantartása és szaporítása
Vírusok szaporítása és vakcinaelőállítási munkafolyamatok
Citotoxicitási és biológiai vizsgálati alkalmazások
Transzfekciós és fehérjeexpressziós vizsgálatok
Alapkutatás a sejtbiológiában és a molekuláris biológiában
Az optimális sejtnövekedés érdekében az EMEM-et általában 5–10 %-os borjúszérummal (FBS) egészítik ki, és a sejtvonal függvényében nem esszenciális aminosavakkal (NEAA) és antibiotikumokkal, például penicillinnel/streptomicinnel.
Kezelés és tárolás
A bontatlan palackot +2 °C és +8 °C között, fénytől védve tárolja. A felnyitás után aszeptikus körülmények között használja. Az oldatban lévő L-glutamin fokozatosan lebomlik – a legjobb teljesítmény érdekében javasoljuk, hogy a tápközeget a felnyitástól számított 4 héten belül használja fel, vagy ha hosszabb ideig tárolja, használat előtt friss L-glutaminnal egészítse ki. A tápközeget hagyja 37 °C-ra melegedni, mielőtt a sejtekhez adagolja.
Minőség
Szigorú minőségi előírások szerint gyártva. Minden tétel sterilitását, pH-értékét, ozmolalitását és endotoxin-szintjét ellenőrizzük, hogy biztosítsuk az állandó teljesítményt a sejtkultúra-alkalmazásokban.
Termékjellemzők
Műszaki adatok
Részlet
TerméktípusMEM
TermékkategóriaSejtkultúra-táptalajok
FormátumFolyékony
SterilIgen
Méret500 ml
L-glutaminL-glutaminnal (2 mM)
GlükózGlükózzal (1,0 g/l)
Nátrium-hidrogén-karbonátNaHCO3-mal (2,2 g/l)
HEPESHEPES nélkül
Nátrium-piruvátNátrium-piruvát nélkül
FenolvörösFenolvörössel
SóoldatEarle kiegyensúlyozott sóoldat (EBSS)
pH7,0 – 7,6
Endotoxin-tartalomNem meghatározva
Tárolás+2 °C és +8 °C között
Összetétel (literenkénti összetétel)
Összetevő
Koncentráció (mg/l)
Szervetlen sók
Kalcium-klorid · 2H2O265,00
Magnézium-szulfát97,72
Kálium-klorid400,00
Nátrium-klorid6 800,00
Nátrium-dihidrogén-foszfát, vízmentes122,00
Nátrium-hidrogén-karbonát (NaHCO3)2 200,00
Aminosavak
L-arginin · HCl126,00
L-cisztin · 2HCl31,30
L-glutamin292,00
L-hisztidin · HCl · H2O42,00
L-izoleucin52,00
L-leucin52,00
L-lizin · HCl72,50
L-metionin15,00
L-fenilalanin32,00
L-treonin48,00
L-triptofán10,00
L-tirozin · 2Na · 2H2O51,90
L-valin46,00
Vitaminok
D-kalcium-pantotenát1,00
Kolin-klorid1,00
Folsav1,00
mio-inozitol2,00
Nikotinamid1,00
Piridoxál · HCl1,00
Riboflavin0,10
Tiamin · HCl1,00
Egyéb összetevők
D(+)-glükóz1 000,00
Fenolvörös10,00

A Freeze Medium CM-1 fő jellemzői a következők:
Széles körű kompatibilitás: Hatékony a sejttípusok széles skálájához, beleértve a primer sejteket, őssejteket és a létrehozott sejtvonalakat.
Magas életképesség: Optimalizált a felolvasztás utáni sejtregeneráció és életképesség maximalizálására, biztosítva a megbízható kísérleti eredményeket.
Felhasználásra kész: Kényelmesen előkészítve és sterilizálva az azonnali alkalmazáshoz, csökkentve az előkészítési időt és a szennyeződés kockázatát.
Fokozott stabilitás: Fenntartja a konzisztens teljesítményt standard kriokonzerválási körülmények között, biztosítva a reprodukálható eredményeket.
Hosszú eltarthatósági idő: A CM-1 egy szérumtartalmú, azonnal felhasználható kriokonzerváló közeg, amely hűtőszekrényben akár egy évig is tárolható.
A CM-1 használata sejtek fagyasztásához
A CM-1 adherens és szuszpenziós sejtek fagyasztására való használatához kövesse az alábbi lépéseket
Adhéziós sejtek esetében mossa meg és válassza le őket a tenyésztési szubsztrátról. Szuszpenziós sejtek esetében közvetlenül a következő lépéssel folytassa.
Számolja meg a sejteket, hogy biztosítsa a megfelelő koncentrációt.
Centrifugáljuk a sejteket a pelletáláshoz, majd szuszpendáljuk újra CM-1 fagyasztási tápfolyadékban.
A reszuszpendált sejteket helyezze át a kriovialis üvegekbe.
Használjon lassú fagyasztási módszert, mielőtt a sejteket hosszú távú tárolásra szállítja
Módszer
Leírás
Lépések
❄️
Kézi fagyasztás
Lépésről lépésre történő módszer, amely a sejtek életképességének biztosítása érdekében a hőmérséklet fokozatos csökkentésével jár
1️⃣ A sejteket fagyasztóközegben 40 percre 4 °C-os fagyasztóba helyezzük.
2️⃣ Tegyük át -80°C-os fagyasztóba 24 órára.
3️⃣ A sejteket folyékony nitrogénben tároljuk a hosszú távú megőrzés érdekében
❄️
Mr. Frosty használata
Kényelmes eszköz, amely lehetővé teszi a szabályozott fagyasztási sebességet elektromos áram nélkül
1️⃣ Készítse elő a sejteket fagyasztóközeggel ellátott kriovialokban.
2️⃣ Helyezze a krioüvegeket a Mr. Frosty tartályba.
3️⃣ Tárolja -80°C-on 24 órán keresztül, mielőtt átrakja folyékony nitrogénbe
❄️
Szabályozott sebességű fagyasztó
A Thermo Fisher vagy más gyártók nagy pontosságú fagyasztója, amelyet szabályozott hőmérsékletcsökkentésre terveztek
1️⃣ Programozza a készüléket a hőmérséklet fokozatos csökkentésére.
2️⃣ Helyezze az előkészített sejteket a fagyasztóba.
3️⃣ A fagyasztási ciklus után helyezze át a sejteket folyékony nitrogénbe
Tárolja a kriovialisokat -130°C alatti hőmérsékleten vagy folyékony nitrogénben a hosszú távú megőrzés érdekében.
Összetevők
FBS-t, DMSO-t, glükózt, sókat tartalmaz
Pufferkapacitás: pH = 7,2 és 7,6 között
A Cytion's Freeze Medium CM-1 megbízható megoldást kínál a kriokonzerváláshoz, biztosítva a sejtek magas életképességét és funkcionalitását a kiolvasztás után a kutatási alkalmazások széles skálájához.

- és csirkemájsejtek tenyésztését, különösen csökkentett szérumtartalmú körülmények között.
A Ham's F-12K (Kaighn's) médiumot gondosan összeállítottuk a sejttenyésztési feltételek optimalizálása érdekében. Dúsított összetétellel rendelkezik, amely megemelt szintet biztosít az olyan esszenciális összetevőkből, mint az aminosavak és a nátrium-piruvát, valamint további elemekből, köztük putreszcinből, timidinből, hipoxantinból és cinkből. Ezek a kiegészítések lehetővé teszik a kutatók számára, hogy a közeget minimális szérummal vagy meghatározott komponensekkel egészítsék ki a specifikus sejttípusok számára, megkönnyítve a pontos kísérleti feltételeket.
Figyelemre méltó, hogy a Ham's F-12K (Kaighn's) Medium nem tartalmaz fehérjéket vagy növekedési faktorokat. Következésképpen gyakran szükséges a növekedési faktorokkal és magzati szarvasmarha szérummal (FBS) való kiegészítés, ami lehetővé teszi a kutatók számára, hogy a médiumot az adott sejtvonalak követelményeihez igazítsák. Az optimális teljesítmény érdekében az FBS koncentrációját gondosan optimalizálni kell az egyes sejtvonalakhoz, biztosítva az optimális növekedést és funkcionalitást.
A fiziológiás pH fenntartása érdekében a Ham's F-12K (Kaighn's) médium nátrium-bikarbonát pufferrendszert alkalmaz (2,5 g/l), ami a tenyésztés során 5-10%-os, ellenőrzött CO2-környezetet tesz szükségessé. Ez biztosítja, hogy a táptalaj pH-ja a sejtek növekedése és életképessége szempontjából ideális tartományban maradjon
Minőségellenőrzés
pH = 7,2 +/
- 0,02 20-25°C-on.
Minden egyes tétel sterilitást, valamint mikoplazma
- és baktériummentességet vizsgáltak.
Karbantartás
Hűtve, +2°C és +8°C között, sötétben tárolva. A fagyasztás, valamint a +37°C-ig történő felmelegítés minimálisra csökkenti a termék minőségét.
Ne melegítse a táptalajt 37°C fölé, és ne használjon ellenőrizhetetlen hőforrásokat (pl. mikrohullámú készülékeket).
Ha a táptalajnak csak egy részét kívánja felhasználni, vegye ki ezt a mennyiséget az üvegből, és melegítse fel szobahőmérsékleten.
Az eltarthatósági idő a gyártási időponttól számított 8 hét bármelyik táptalaj esetében, kivéve az alap táptalajt.
Összetétel
Összetevők
mg/l
Szervetlen sók
Kalcium-klorid x 2H2O
135,24
Réz(II)szulfát x 5H2O
0,00
Vas(II)szulfát x 7H2O
0,83
Magnézium-klorid x 6H2O
105,72
Magnézium-szulfát x 7H2O
394,49
Kálium-klorid
283,29
Kálium-dihidrogén-foszfát
58,52
Nátrium-klorid
7597,20
di-nátrium-hidrogén-foszfátvízmentes
115,02
Cink-szulfát x 7H2O
0,14
Egyéb összetevők
D(+)-Glükóz vízmentes
1260,00
Hipoxantin
4,08
DL-α-Liponsav
0,21
Fenolvörös
3,00
Putreszcin x 2HCl
0,32
Nátrium-piruvát
220,00
NaHCO3
2500,00
Timidin
0,73
Aminosavak
L-Alanin
17,82
L-arginin x HCl
421,40
L-aszparagin x H2O
30,02
L-aszparaginsav
26,62
L-cisztein x HCl x H2O
70,24
L-Glutamin
292,20
L-Glutaminsav
29,42
Glicin
15,01
L-Hisztidin x HCl x H2O
41,92
L-izoleucin
7,87
L-Leucin
26,24
L-lizin x HCl
73,04
L-metionin
8,95
L-fenilalanin
9,91
L-prolin
69,06
L-szerin
21,02
L-treonin
23,82
L-triptofán
4,08
L-tirozin
10,87
L-Valin
23,42
Vitaminok
D(+)-Biotin
0,07
D-kalcium-pantotenát
0,48
Kolin-klorid
13,96
Folsav
1,32
myo-Inozitol
18,02
Nikotinamid
0,04
Piridoxin x HCl
0,06
Riboflavin
0,04
Tiamin x HCl
0,34
B12-vitamin
1,36

A foszfátpufferelt sóoldat (PBS) a biológiai és kémiai kutatásokban széles körben használt pufferoldat. Fontos szerepet játszik a pH-egyensúly és az ozmolaritás fenntartásában a különböző kísérleti eljárások során, beleértve a szövetfeldolgozást és a sejttenyésztést. PBS-oldatunkat nagy gondossággal, nagy tisztaságú összetevőkkel állítjuk össze, hogy minden kísérletben biztosítsuk a stabilitást és a megbízhatóságot. PBS-ünk ozmolaritása és ionkoncentrációja szorosan utánozza az emberi szervezetben uralkodó értékeket, így izotóniás és a legtöbb sejtre nem mérgező.
PBS-oldatunk összetétele
A PBS oldatunk ultratisztaságú foszfátpufferek és sóoldatok pH-értékre beállított keveréke. Az 1X munkakoncentrációban a következőket tartalmazza:
8000 mg/l nátrium-klorid (NaCl)
200 mg/l kálium-klorid (KCl)
1150 mg/L nátrium-foszfát kétbázisú vízmentes (Na2HPO4)
200 mg/L Kálium-foszfát monobázikus vízmentes (KH2PO4)
Ez az összetétel optimális pH
- és ionegyensúlyt biztosít, amely a biológiai alkalmazások széles köréhez alkalmas.
A PBS-oldatunk alkalmazásai
PBS-oldatunk ideális a biológiai kutatások különböző alkalmazásaihoz. Izotóniás és nem toxikus tulajdonságai alkalmassá teszik az anyagok hígítására és a sejttartályok öblítésére. Az EDTA-t tartalmazó PBS-oldatok hatékonyan oldják a megtapadt és csomósodott sejteket. Kétértékű fémeket, például cinket azonban nem szabad a PBS-hez adni, mivel ez csapadékképződést okozhat. Ilyen esetekben a Good pufferek használata ajánlott. Ezenkívül a PBS-oldatunk elfogadható alternatívája a vírusszállító közegnek az RNS-vírusok, köztük a SARS-CoV-2 szállítására és tárolására.
Minőségellenőrzés
Steril szűrt
Tárolás és eltarthatósági idő
Tárolja +2°C és +25°C között, fénytől védve.
Felbontás után 2°C és 25°C között tárolja, és 24 hónapon belül használja fel.
Szállítási feltételek
Környezeti hőmérséklet
Karbantartás
Hűtve, +2°C és +8°C között, sötétben tárolja. Kerülje a fagyasztást és a gyakori melegítést +37°C-ra, mivel ez csökkenti a termék minőségét.
Ne melegítse a közeget 37°C fölé, és ne használjon ellenőrizetlen hőforrásokat, például mikrohullámú készülékeket.
Ha a táptalajnak csak egy részét kívánja felhasználni, vegye ki a szükséges mennyiséget, és felhasználás előtt melegítse fel szobahőmérsékletűre.
Összetétel
Kategória
Összetevők
Koncentráció (mg/l)
Sók
Kálium-klorid
200
Kálium-foszfát monobázikus vízmentes
200
Nátrium-klorid
8000
Nátrium-foszfát kétbázisú, vízmentes
1150

Az RPMI 1640 médiumot eredetileg humán leukémiás sejtek szuszpenziós és monolayer kultúrákban történő növekedésének támogatására tervezték, de a kutatók és a kereskedelmi beszállítók módosításai révén azóta az emlőssejtek sokféle fajtájához alkalmassá vált. Kivételesen kompatibilis az olyan sejtvonalakkal, mint a HeLa, Jurkat, MCF-7, PC12, PBMC, asztrociták és karcinómák.
Az RPMI 1640 médium egyedi összetétele miatt különbözik más sejttenyésztő médiumoktól. Jelentős mennyiségű foszfátot, aminosavakat és vitaminokat tartalmaz. Különösen biotint, B12-vitamint és PABA-t tartalmaz, amelyek hiányoznak az Eagle's Minimal Essential Mediumból vagy a Dulbecco's Modified Eagle Mediumból. Továbbá az RPMI 1640 Medium jelentősen megemelkedett koncentrációban tartalmaz inozitol és kolin vitaminokat. Nem tartalmaz azonban fehérjéket, lipideket vagy növekedési faktorokat. Következésképpen a sejtnövekedés optimális feltételeinek biztosításához általában 10%-os magzati szarvasmarha-szérummal (FBS) való kiegészítésre van szükség.
Az RPMI 1640 médium pufferrendszere nátrium-bikarbonátra támaszkodik, és 5-10%-os CO2-környezetet igényel a fiziológiailag megfelelő pH fenntartásához. A glutation redukálószer beépítése tovább különbözteti meg ezt a táptalajt a többitől.
Minőségellenőrzés
Steril szűrt
Tárolás és eltarthatóság
+2°C és +8°C között, fénytől védve tárolja.
Felbontás után 4°C-on tárolja, és 6-8 héten belül használja fel.
Szállítási feltételek
Környezeti hőmérséklet
Karbantartás
Hűtve, +2°C és +8°C között, sötétben tárolja. Kerülje a fagyasztást és a gyakori melegítést +37°C-ra, mivel ez csökkenti a termék minőségét.
Ne melegítse a közeget 37°C fölé, és ne használjon ellenőrizetlen hőforrásokat, például mikrohullámú készülékeket.
Ha a táptalajnak csak egy részét kívánja felhasználni, vegye ki a szükséges mennyiséget, és felhasználás előtt melegítse fel szobahőmérsékletűre.
Összetétel
Kategória
Összetevők
Koncentráció (mg/l)
Aminosavak
Glicin
10.00
L-Alanil-L-Glutamin
434.40
L-arginin
200.00
L-aszparaginH2O
56.82
L-aszparaginsav
20.00
L-cisztin 2HCl
65.20
L-Glutaminsav
20.00
L-Hisztidin HClH2O
20.27
L-hidroxi-L-prolin
20.00
L-izoleucin
50.00
L-Leucin
50.00
L-lizin HCl
40.00
L-metionin
15.00
L-fenilalanin
15.00
L-Prolin
20.00
L-szerin
30.00
L-treonin
20.00
L-triptofán
5.00
L-Tirozin 2Na 2H2O
28.83
L-Valin
20.00
Vitaminok
p-Amino benzoesav
1.00
D-Biotin
0.20
Kolinklorid
3.00
D-kalcium-pantotenát
0.25
Folsav
1.00
myo-Inozitol
35.00
Nikotinamid
1.00
Piridoxin HCl
1.00
Riboflavin
0.20
Tiamin HCl
1.00
B12-vitamin
0.005
Szervetlen sók
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Egyéb összetevők
D-Glükóz
2000.00
L-Glutation redukált
1.00
Fenolvörös nátriumsó
5.30
, w: 3,1 g/L glükóz, w: 2,5 mM L-Glutamin, w: 15 mM HEPES, w: 0,5 mM nátrium-piruvát, w: 1,2 g/L NaHCO3")
Ez az egyedülálló készítmény a Dulbecco's Modified Eagle Medium (DMEM) és a Ham's F-12 (Ham's Nutrient Mixture F-12) tápközeget pontos 1:1 arányban ötvözi. Az L-glutamin hozzáadása tovább javítja összetételét.
Az Eagle's Minimal Essential Medium (EMEM) alapú DMEM elődjéhez képest megnövelt koncentrációban tartalmaz aminosavakat és vitaminokat. Ezzel szemben a Ham's F-12 a Ham's F-10 táptalajon alapul, és kiegészítő esszenciális komponenseket biztosít.
Az optimális sejtnövekedés támogatása érdekében általános gyakorlat a DMEM:Ham's F12 kiegészítése FBS-sel, jellemzően 5–10%-os koncentrációban. Ez a kiegészítés szükséges, mivel a táptalajból hiányoznak a sejtek fejlődéséhez elengedhetetlen növekedési hormonok, lipidek és fehérjék.
A DMEM:Ham's F12 pH-puffer rendszert tartalmaz, és gyakran kiegészítik fenolvörössel, egy pH-indikátorral. A DMEM:Ham's F12-ben, vagy bármelyik bikarbonát-puffer rendszert alkalmazó táptalajban tenyésztett sejteknek 5–10%-os szabályozott CO2-környezetre van szükségük a megfelelő pH-szint fenntartásához.
Minőség-ellenőrzés
Steril szűrt
Tárolás és eltarthatóság
+2 °C és +8 °C között, fénytől védve tárolandó.
A csomag felbontása után 4 °C-on tárolandó, és 6–8 héten belül fel kell használni.
Szállítási feltételek
Környezeti hőmérséklet
Tárolás
Sötét helyen, +2 °C és +8 °C között hűtőszekrényben tárolandó. Kerülje a fagyást és a gyakori +37 °C-ra történő felmelegítést, mivel ez rontja a termék minőségét.
Ne melegítse a táptalajt 37 °C fölé, és ne használjon ellenőrizhetetlen hőforrásokat, például mikrohullámú sütőt.
Ha a táptalaj csak egy részét használja fel, vegye ki a szükséges mennyiséget, és használat előtt melegítse szobahőmérsékletre.
Összetétel
Kategória
Összetevők
Koncentráció (mg/l)
Aminosavak
Glicin
18,75
L-alanin
4,45
L-arginin-HCl
147,50
L-aszparagin-H₂O
7,50
L-aszparaginsav
6,65
L-cisztein-HCl-H₂O
17,56
L-cisztin-2-HCl
31,29
L-glutaminsav
7,35
L-glutamin
365,00
L-hisztidin-HCl-H₂O
31,48
L-izoleucin
54,47
L-leucin
59,05
L-lizin-HCl
91,25
L-metionin
17,24
L-fenilalanin
35,48
L-prolin
17,25
L-szerin
26,25
L-treonin
53,45
L-triptofán
9,02
L-tirozin 2 Na 2 H2O
55,79
L-valin
52,85
Vitaminok
D-biotin
0,0035
Kolin-klorid
8,98
D-kalcium-pantotenát
2,24
Folsav
2,66
mio-inozitol
12,60
Nikotinamid
2,02
Piridoxin-HCl
0,031
Piridoxál-HCl
2,00
Riboflavin
0,219
Tiamin-HCl
2,17
B12-vitamin
0,68
Szervetlen sók
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO3)3 9 H2O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Egyéb összetevők
D-glükóz
3151,00
Hipoxantin
2,40
HEPES
3574,50
Linolsav
0,042
Liponsav
0,105
Fenolvörös nátriumsó
8,63
Putrescin-2-HCl
0,081
Nátrium-piruvát
55,00
Timidin
0,365
, w: 3,0 g/L glükóz, w: stabil glutamin, w: 2,0 mM nátrium-piruvát, w: 2,2 g/L NaHCO3")
- 0,02 20-25 °C-on. Minden tétel sterilitását, valamint mikoplazma és baktériumok hiányát tesztelték. Tárolás Hűvös helyen, +2 °C és +8 °C között tárolandó. A fagyasztás és a +37 °C-ra történő felmelegítés rontja a termék minőségét. Ne melegítse a táptalajt 37 °C fölé, és ne használjon ellenőrizhetetlen hőforrásokat (pl. mikrohullámú sütőt). Ha csak a táptalaj egy részét kívánja felhasználni, vegye ki azt a palackból, és melegítse szobahőmérsékletre. Az alapközeg kivételével minden közeg szavatossági ideje a felnyitástól számított 6–8 hét. Összetétel Összetevők mg/L Szervetlen sókKalcium-klorid x 2H2O132,00 Magnézium-szulfát97,67 Kálium-klorid400 Nátrium-klorid6460 Dinátrium-hidrogén-foszfát (vízmentes)504,00 Egyéb összetevőkD(+)-glükóz (vízmentes)3 000,00 Glutation (redukált)0,5 Húspepton600,00 Fenol-vörös nátriumsó11 AminosavakL-alanin13,36 L-arginin x HCl42,14 L-aszparagin x H2O45,03 L-aszparaginsav19,97 L-cisztein x HCl x H2O31,75 L-glutamin (stabil)219,15 L-glutaminsav22,07 Glicin7,51 L-hisztidin x HCl x H2O20,96 L-hidroxiprolin19,67 L-izoleucin39,36 L-leucin39,36 L-lizin x HCl36,54 L-metionin14,92 L-fenilalanin16,52 L-prolin17,27 L-szerine26,28 L-treonin17,87 L-triptofán3,06 L-tirozin-dinátrium-só26,10 L-valin17,57 Vitaminokp-Aminobenzoesav1,0 Aszkorbinsav0,56 D(+)-biotin0,20 D-kalcium-pantotenát0,20 Kolin-klorid5,0 Folsav10 mio-inozitol36 Nikotinamid0,5 Nikotinsav0,5 Piridoxál-HCl0,50 Piridoxin-HCl0,50 Riboflavin0,2 Tiamin-HCl0,2 B12-vitamin2,00

A 199-es médium számos alkalmazási lehetőséget kínál a területen. Hatékonyan képes fenntartani a kumulus-oocita komplexet (COC) és támogatni az oociták in vitro érését. Ezenkívül a német holstein tehenekből történő petesejtgyűjtés során az aspirációs vonalak öblítésénél is alkalmazzák. A Medium 199 továbbá kiváló táptalajként szolgál a patkányokból származó szív endotélsejtek tenyésztéséhez. Ezek az alkalmazások bizonyítják a Medium 199 sokoldalúságát és alkalmazkodóképességét a különböző kísérleti igényekhez.
Történet
A Medium 199 kifejlesztése az 1950-es években jelentős előrelépést jelentett a szövettenyésztő közegek terén. Bevezetése előtt számos tenyésztőközeg állati eredetű termékekre és szövetkivonatokra támaszkodott. Morgan és munkatársai azonban forradalmasították a területet azzal, hogy egy teljesen meghatározott tápanyagforrást fogalmaztak meg a sejtkultúrák számára. A vitaminok, aminosavak és egyéb tényezők különböző kombinációival végzett kísérleteik során felfedezték a Medium 199 kivételes növekedésserkentő tulajdonságait.
Minőségellenőrzés
pH = 7,2 +/
- 0,02 20-25°C-on.
Minden egyes tétel sterilitását, valamint mikoplazma
- és baktériummentességét vizsgálták.
Karbantartás
Hűtve, +2°C és +8°C között, sötétben tárolva. A fagyasztás és a melegítés +37°C-ig minimalizálja a termék minőségét.
Ne melegítse a táptalajt 37° C-nál magasabbra, és ne használjon ellenőrizhetetlen hőforrásokat (pl. mikrohullámú készülékeket).
Ha a táptalajnak csak egy részét kívánja felhasználni, vegye ki ezt a mennyiséget az üvegből, és melegítse fel szobahőmérsékleten.
Az eltarthatósági idő az alapközeg kivételével bármelyik táptalaj esetében a gyártás dátumától számított 8 hét.
Összetétel
Összetevők
mg/l
Szervetlen sók
Kalcium-klorid x 2H2O
264,92
Vas(III)nitrát x 9H2O
0,72
Magnézium-szulfát
97,67
Kálium-klorid
400,00
Nátrium-acetát x 3H2O
82,95
Nátrium-klorid
6,800.00
Nátrium-dihidrogén-foszfát x H2O
140,00
Egyéb összetevők
Adenin-szulfát
10,00
AMP
0,20
ATP
1,00
Koleszterin
0,20
2'-Deoxiribóz
0,50
D(+)-Glükóz vízmentes
1,000.00
Glutation (vörös)
0,05
Guanin x HCl
0,30
Hipoxantin
0,30
Fenolvörös
10,00
D-Ribóz
0,50
Thymin
0,30
Tween 80
4,90
Uracil
0,30
Xantin
0,30
NaHCO3
2,200.00
Aminosavak
L-Alanin
25,00
L-arginin x HCl
70,00
L-aszparaginsav
30,00
L-cisztein x HCl x H2O
0,10
L-cisztin
20,00
L-Glutamin stabil
149,00
L-Glutaminsav
67,00
Glicin
50,00
L-Hisztidin x HCl x H2O
21,88
L-hidroxiprolin
10,00
L-izoleucin
20,00
L-Leucin
60,00
L-lizin x HCl
70,00
L-metionin
15,00
L-fenilalanin
25,00
L-prolin
40,00
L-szerin
25,00
L-treonin
30,00
L-triptofán
10,00
L-tirozin
40,00
L-Valin
25,00
Vitaminok
4-Amino benzoesav
0,05
Aszkorbinsav
0,05
D(+)-Biotin
0,01
Kalciferol
0,10
D-kalcium-pantotenát
0,01
Kolin-klorid
0,50
Folsav
0,01
myo-Inozitol
0,05
Menadion
0,01
Nikotinsav
0.025
Nikotinamid
0.025
Piridoxal x HCl
0.025
Piridoxol x HCl
0.025
Riboflavin
0,01
DL-α-tokoferol-foszfát-dinátrium só
0,01
Tiamin x HCl
0,01
A-vitamin-acetát
0,14

Az IMDM jól alkalmazható gyorsan proliferáló, nagy sűrűségű sejtkultúrákhoz, beleértve a Jurkat, COS-7 és makrofág sejteket. Az IMDM különböző sejttenyésztési alkalmazásokhoz rendelkezésre álló különböző módosításai megtalálhatók a médiaválasztó eszköz segítségével. A folyékony médiumok alapvető tápanyagokat biztosítanak minden sejttenyésztési alkalmazáshoz. Minden egyes kiváló minőségű sejttenyésztő táptalajunkat az eredetileg közzétett receptúra vagy a táptalaj konzisztens teljesítményéhez és stabilitásához szükséges módosítások szerint állítjuk elő.
IMDM vs. DMEM
Az IMDM vas-nitrát helyett kálium-nitrátot, valamint HEPES-t és nátrium-piruvátot tartalmaz. Az IMDM-ben lévő további összetevők miatt az IMDM alkalmasabb speciális sejttípusok és speciális alkalmazások számára, mint a DMEM.
IMDM vs. RPMI
Az IMDM és az RPMI eltérő összetételű, ami a PMA/ionomicin stimuláció szempontjából fontos lehet. Az egyik jelentős különbség a Ca2+ koncentrációja. Míg az RPMI 0,42 mM Ca2+-t tartalmaz, addig az IMDM 1,49 mM-ot.
Minőségellenőrzés
pH = 7,2 +/
- 0,02 20-25°C-on.
Minden egyes tétel sterilitási, valamint mikoplazma
- és baktériummentességi vizsgálaton esett át.
Karbantartás
Hűtve, +2°C és +8°C között, sötétben tárolva. A fagyasztás és a melegítés +37°C-ig minimalizálja a termék minőségét.
Ne melegítse a táptalajt 37° C-nál magasabbra, és ne használjon ellenőrizhetetlen hőforrásokat (pl. mikrohullámú készülékeket).
Ha a táptalajnak csak egy részét kívánja felhasználni, vegye ki ezt a mennyiséget az üvegből, és melegítse fel szobahőmérsékleten.
Az alapközeg kivételével bármelyik táptalaj eltarthatósági ideje a gyártás dátumától számított 8 hét.
Összetétel
Összetevők
mg/l
Szervetlen sók
Kalcium-klorid x 2 H2O
219,00
Kálium-klorid
330,00
Kálium-nitrát
0.076
Magnézium-szulfát vízmentes
97,73
Nátrium-klorid
4,505.00
Nátrium-dihidrogén-foszfát vízmentes
109,00
Nátrium-szelenit
0,02
Egyéb összetevők
D(+)-Glükóz vízmentes
4,500.00
HEPES
5,958.00
Nátrium-piruvát
110,00
Fenolvörös
15,00
Aminosavak
L-alanin
25,00
L-arginin x HCl
84,00
L-aszparagin x H2O
25,00
L-aszparaginsav
30,00
L-cisztin x 2HCl
91,24
L-Glutamin
584,00
L-Glutaminsav
75,00
Glicin
30,00
L-Hisztidin x HCl x H2O
42,00
L-izoleucin
104,80
L-Leucin
104,80
L-lizin x HCl
146,20
L-metionin
30,00
L-fenilalanin
66,00
L-prolin
40,00
L-szerin
42,00
L-treonin
95,20
L-triptofán
16,00
L-tirozin x 2Na
104,20
L-Valin
93,60
Vitaminok
D(+)-Biotin
0.013
D-kalcium-pantotenát
4,00
Kolin-klorid
4,00
Folsav
4,00
myo-Inozitol
7,20
Sejtkultúra-táptalajok: Áttekintés
Az élettudományok területén az egyik legfontosabb módszer a sejtkultúra. A „sejtkultúra” kifejezés alatt azt értjük, hogy sejteket, szöveteket vagy szerveket veszünk ki egy állatból vagy növényből, majd azokat olyan mesterséges környezetbe ültetjük át, amely kedvező a túlélésükhöz és/vagy növekedésükhöz. Az optimális sejtfejlődés alapvető környezeti feltételei a szabályozott hőmérséklet, a sejtek tapadásához szükséges szubsztrátum, a megfelelő tenyészközeg, valamint az optimális pH-értéket és ozmolalitást fenntartó inkubátor. A sejteknek ezekre a feltételekre van szükségük ahhoz, hogy teljes potenciáljukat kihasználva növekedhessenek.
A megfelelő tenyészközeg kiválasztása az in vitro tenyésztéshez a sejtkultúra legkritikusabb és egyben legfontosabb szakasza. A tenyésztőközeg – más néven tenyésztőtáp – olyan folyadék vagy gél, amelyet úgy állítottak össze, hogy elősegítse a szervezetek fejlődését mikroszkopikus, sejt- vagy növényi szinten. A sejtek tenyésztéséhez használt közeg gyakran megfelelő mennyiségű energiát és olyan anyagokat tartalmaz, amelyek szabályozzák a sejtciklust. A tenyésztőközeg fő összetevői közé tartoznak az aminosavak, a vitaminok, a szervetlen sók, a glükóz és a szérum. A szérumot azért adják a közeghez, mert növekedési faktorok, hormonok és tapadási faktorok forrásaként szolgál. A tápanyagok biztosítása mellett a közeg hozzájárul a pH-érték és az ozmolalitás szintjének fenntartásához is.
A sejtkultúrában használt táptalajok típusai
Mind az emberi, mind az állati sejtek tenyészthetők mesterséges vagy szintetikus táptalajon, illetve természetes elemekkel kiegészített, teljesen természetes táptalajon. Az alábbiakban áttekintést adunk a jelenleg rendelkezésre álló különböző táptalajtípusokról.
Természetes táptalajok
A természetes táptalajokban kizárólag természetes állapotban előforduló biológiai folyadékok találhatók. A természetes táptalajok nagyon hasznosak és könnyen alkalmazhatók számos különböző állati sejttípus tenyésztéséhez. A természetes táptalajokat alkotó pontos összetevők ismeretének hiánya az elsődleges tényező, amely hozzájárul a természetes táptalajok használatával kapott eredmények alacsony megismételhetőségéhez.
Mesterséges táptalajok
A mesterséges vagy szintetikus táptalajok előállításához tápanyagok (szerves és szervetlen egyaránt), szérumfehérjék, szénhidrátok, kofaktorok, vitaminok és sók, valamint O2 és CO2 gázfázisok hozzáadása szükséges [1].
Különböző típusú mesterséges táptalajokat fejlesztettek ki a következő funkciók közül egy vagy több teljesítése érdekében: 1) Azonnali túlélés (kiegyensúlyozott sóoldat pontos pH-értékkel és ozmotikus nyomással). 2) Hosszabb idejű túlélés (kiegyensúlyozott sóoldat, amelyet különböző összetételű szerves vegyi anyagokkal és/vagy szérummal egészítenek ki). 3) Határidő nélküli fejlődés. 4) Speciális funkciók.
A mesterséges táptalajok négy különböző osztályba sorolhatók:
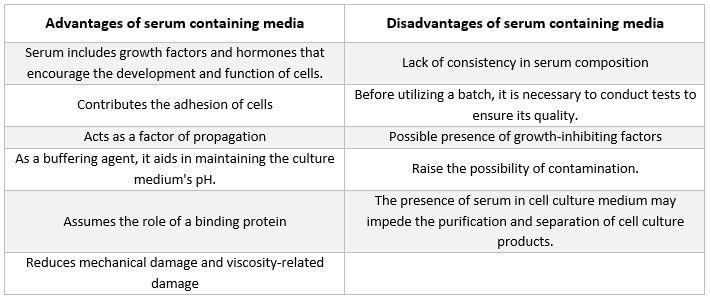
Szérumtartalmú táptalajok
Az állati sejtek tenyésztéséhez használt táptalajokban leggyakrabban előforduló kiegészítő a borjúszérum. Ezt olcsó kiegészítőként adják a tenyésztőtáptalajhoz a lehető legjobb növekedési feltételek elérése érdekében. A szérum nemcsak a labilis vagy vízben oldhatatlan tápanyagok, hormonok és növekedési faktorok, proteázgátlók és egyéb anyagok szállítójaként vagy kelátképzőjeként működik, hanem a káros molekulákat is megköti és semlegesíti.
Szérummentes táptalaj
A szérum jelenléte a táptalajban számos hátránnyal jár, és az immunológiai kutatásokban súlyos értelmezési hibákhoz vezethet [2, 3]. Számos különböző szérummentes táptalajt fejlesztettek ki [4, 5]. Ezeket a táptalajokat általában egy adott sejttípus tenyésztésének támogatására fejlesztették ki, például a Thermo Fisher Scientific Knockout Serum Replacement és Knockout DMEM táptalajait, valamint a Stem Cell Technologies mTESR táptalaját [6] az őssejtek számára [7].
Ezenkívül ezek a táptalajok meghatározott mennyiségű tisztított növekedési faktort, lipoproteint és egyéb fehérjét tartalmaznak, amelyeket egyébként általában a szérum biztosít [8]. Ezeket a táptalajokat gyakran „meghatározott összetételű tenyésztőtáptalajoknak” nevezik, mivel összetevőik jól ismertek.
Kémiailag meghatározott táptalajok
Ezek a táptalajok olyan ultratiszta szervetlen és szerves összetevőket tartalmaznak, amelyek semmiféle szennyeződéstől mentesek. Tartalmazhatnak továbbá tiszta fehérje-kiegészítőket is, például növekedési faktorokat.
A baktériumok vagy élesztők genetikai módosítása, valamint bizonyos zsírsavak, vitaminok, koleszterin és aminosavak hozzáadása eredményeként keletkeznek ezek az összetevők [9].
Fehérje-mentes táptalajok
A fehérje-mentes táptalajok olyanok, amelyek egyáltalán nem tartalmaznak fehérjét, hanem kizárólag nem fehérje jellegű elemeket. A szérummal dúsított táptalajokhoz képest a fehérje-mentes táptalajok használata elősegíti a sejtek fokozottabb szaporodását és a fehérje-expressziót, valamint megkönnyíti a későbbi feldolgozási folyamatokban keletkező termékek tisztítását [10–12]. Az olyan készítményekben, mint az MEM és az RPMI-1640, nincs fehérje. Szükség esetén azonban fehérje-kiegészítőt is adagolhatnak.
Tenyésztőközeg és alapvető összetevői
A kereskedelmi forgalomban kapható tenyésztőközeg por vagy folyadék formájában szerezhető be, és gyakran tartalmaz különféle tápanyagokat, például aminosavakat, glükózt, sókat, vitaminokat és egyéb étrend-kiegészítőket.
Ezen összetevők iránti igény minden sejtvonal esetében eltérő, és ezek a különbségek magyarázzák a táptalajok széles körű összetételét. Minden összetevő egy bizonyos funkciót lát el, amelyeket a következő bekezdésekben ismertetünk:
Pufferrendszerek
Az optimális tenyésztési feltételek fenntartása érdekében a pH-értéket szabályozni kell, ami gyakran kétféle pufferrendszer egyikével történik:
Természetes pufferrendszer
A légkörben a CO2/H2CO3 arány megegyezik a tápközegben mért értékkel, ami természetes puffermechanizmust hoz létre. A természetes puffermechanizmus megőrzése érdekében a tenyészeteket 5–10%-os CO2-tartalmú levegőben kell tartani, amit gyakran CO2-inkubátor használatával érnek el. A természetes puffer használatának egyik legnagyobb előnye, hogy olcsó és biztonságos.
HEPES
A HEPES zwitteriont alkalmazó kémiai pufferelés nagyobb pufferelési képességgel rendelkezik a 7,2–7,4 közötti pH-tartományban, és nem igényel szabályozott gázkörnyezetet. Bizonyos sejttípusok esetében a HEPES nagyobb adagja káros lehet. A HEPES-t tartalmazó táptalajok szintén sokkal érzékenyebbek a fluoreszkáló fény fototoxikus hatásaira [13].
Fenolvörös
A pH-indikátorként szolgáló fenolvörös gyakran megtalálható a kereskedelemben kapható tenyésztőközegben, lehetővé téve a pH folyamatos figyelemmel kísérését. A sejtek szaporodásával az általuk termelt metabolitok a pH-érték eltolódását, és ezáltal a tápközeg színének megváltozását idézik elő. A fenolvörös kettős hatást gyakorol a tápközeg színére: savas pH-értéknél sárgává, lúgos pH-értéknél pedig lilává változtatja azt. A sejtkultúrához optimális pH-érték, a 7,4, a tápközeget fluoreszkáló vörös színűvé teszi.
A fenolvörösnek azonban van néhány hátránya: Először is, a fenolvörös képes számos szteroid hormon, elsősorban az ösztrogén hatását szimulálni [14]. Ezért ösztrogénérzékeny sejtek, például emlőszövet vizsgálata során fenolvörösmentes tápközeg használata ajánlott. Számos szérummentes készítményben a fenolvörös jelenléte megzavarja a nátrium–kálium egyensúlyt. A tápközeghez szérum vagy szarvasmarha agyalapi mirigy hormon hozzáadása ellensúlyozhatja ezt a hatást [15]. Harmadszor, a fenolvörös jelenléte akadályozza a detektálást áramlási citometriás kísérletek során.
Szervetlen sók
A szervetlen sókat, például nátrium-, kálium- és kalciumionokat tartalmazó táptalajok elősegítik az ozmotikus egyensúly fenntartását és szabályozzák a membránpotenciált.
Aminosavak
Mivel az aminosavak a fehérjék alapvető alkotóelemei, minden eddig kifejlesztett sejttenyésztő tápközeg elengedhetetlen alkotóelemei. Mivel a sejtek bizonyos aminosavakat nem képesek önállóan előállítani, fontos, hogy a tenyésztőközeg tartalmazza az esszenciális aminosavakat. Ezek szükségesek a sejtek szaporodásához, és jelenlétük koncentrációja határozza meg az elérhető maximális sejtsűrűséget. Különösen fontos az L-glutamin, egy esszenciális aminosav.
Az L-glutamin az anyagcsere másodlagos energiaforrásaként működik, és nitrogént biztosít a NAD, a NADPH és a nukleotidok előállításához. Mivel az L-glutamin egy instabil aminosav, amely idővel olyan formává alakul át, amelyet a sejtek már nem tudnak hasznosítani, ezért azt a tápközegbe kell adagolni.
Ezen felül nem esszenciális aminosavakat is be lehet juttatni a tápközegbe, hogy pótolják a növekedési folyamat során felhasználtakat. A sejtek növekedése felgyorsul, és életképességük javul, ha a tápközeget nem esszenciális aminosavakkal egészítik ki.
Szénhidrátok
A cukor formájában előforduló szénhidrátok jelentik a fő energiaforrást. Számos táptalaj a glükóz és a galaktóz mellett maltózt és fruktózt is tartalmaz.
Fehérjék és peptidek
Az albumin, a transzferrin és a fibronektin a leggyakrabban használt fehérjék és peptidek. Különösen fontosak a szérumot nem tartalmazó táptalajokban. Az albumin, a transzferrin, az aprotinin, a fetuin és a fibronektin a szérumban előforduló fehérjék közé tartoznak, amely gazdag fehérjeforrás.
Az albumin a vérben található fő fehérje, amelynek feladata különböző anyagok – többek között víz, sók, szabad zsírsavak, hormonok és vitaminok – megkötése és szállítása a különböző szervek és sejtek között. Az albumin kémiai anyagokhoz való kötődési képessége miatt hatékonyan alkalmazható a sejtek tenyésztésére szolgáló táptalajból a káros vegyületek eltávolítására.
Az aprotinin védőanyagként szolgál a sejtkultúra-rendszerekben, mivel semleges és savas pH-érték mellett is stabil, valamint ellenáll a magas hőmérsékletnek és a proteolitikus enzimek által okozott lebomlásnak. Képes számos szerinproteáz gátlására, többek között a tripszinre is.
A fetuin egy glikoprotein, amely a felnőtt állatok szérumához képest nagyobb mennyiségben mutatható ki a magzati és újszülött állatok szérumában. Ezen felül szerinproteáz-gátlóként is hat. A fibronektin fehérje a sejtadhézió folyamatának elengedhetetlen alkotóeleme. A transzferrin egy fehérje, amely a vasat szállítja, és felelős a vas sejtmembránokba történő eljuttatásáért.
Zsírsavak és lipidek
A szérum hiányában döntő szerepet játszanak a szérummentes táptalajban.
Vitaminok
Számos vitamin szükséges a sejtek fejlődéséhez és szaporodásához. A sejtek nem képesek megfelelő mennyiségben előállítani a vitaminokat, ezért a szövetkultúrában étrend-kiegészítőként elengedhetetlenek.
A sejtkultúrában a szérum a vitaminok elsődleges forrása; a táptalajokat azonban különféle vitaminokkal is dúsítják, hogy azok egy adott sejttípus számára megfelelőek legyenek. A növekedés serkentésére leggyakrabban a B-csoportba tartozó vitaminokat használják.
Nyomelemek
Az olyan kémiai elemek, mint a réz, a cink, a szelén és a trikarbonsav-köztes termékek, nyomelemek néven ismertek. A nyomelemeket gyakran adják hozzá a szérumot nem tartalmazó táptalajokhoz, hogy pótolják a szérumban általában jelen lévőket. Ezek az elemek fontos kémiai összetevők, amelyek szükségesek az egészséges sejtfejlődéshez. Számos biokémiai reakció – például az enzimaktivitás – bizonyos mikrotápanyagoktól függ.
Tápközeg-kiegészítők
Az egyes sejtvonalak számára javasolt teljes növekedési táptalajhoz olyan kiegészítő összetevőkre van szükség, amelyek nem találhatók meg az alap-táptalajban és a szérumban. Ezek a táplálékkiegészítők támogatják a sejtek növekedését és a megfelelő anyagcserét.
Bár a hormonok, a növekedési faktorok és a jelátviteli molekulák elengedhetetlenek bizonyos sejtvonalak megfelelő szaporodásához, mindig be kell tartani a következő óvintézkedéseket: Mivel a kiegészítők hozzáadása megváltoztathatja a teljes tenyészközeg ozmolalitását, ami gátolhatja a sejtek fejlődését, ezért a kiegészítők hozzáadása után mindig célszerű ellenőrizni az ozmolalitást. A sejtvonalak többségénél az optimális ozmolalitás 260 és 320 mOSM/kg között mozog.
Antibiotikumok
Az antibiotikumokat gyakran alkalmazzák a baktériumok és gombák okozta szennyeződések gátlására [16], bár a sejtnövekedéshez nem elengedhetetlenek. Mivel az antibiotikumok elrejthetik a mikoplazma és az antibiotikum-rezisztens baktériumok által okozott szennyeződéseket, rutinszerű alkalmazásuk nem javasolt a sejtkultúrákban [17, 18].
Ezen felül az antibiotikumok megzavarhatják az érzékeny sejtek anyagcseréjét. Gyakran használják a MilliporeSigma és a Life Technologies által gyártott penicillin–streptomicin kombinációkat. A Plasmocint a TS603, TS516 és BT260 glióma sejtvonalak tenyésztésében alkalmazták [19], és kimutatták, hogy hatékony a mikoplazma-fertőzés eltávolításában (20).
Szérum
A szérumban albuminok, növekedési faktorok és növekedésgátlók egyaránt jelen vannak. A szérum a sejtkultúra-táptalaj egyik legfontosabb alkotóeleme, mivel aminosavakat, fehérjéket, vitaminokat (különösen zsírban oldódó vitaminokat, mint az A-, D-, E- és K-vitamin), szénhidrátokat, lipideket, hormonokat, növekedési faktorokat, ásványi anyagokat és nyomelemeket biztosít.
A tenyésztett sejtek fejlődésének elősegítésére gyakran használnak magzati és borjú származású szérumot. A magzati szérum bőséges növekedési faktor-forrás, és alkalmas sejtklónozásra, valamint érzékeny sejtek tenyésztésére. Csökkent növekedésserkentő képessége miatt a borjúszérumot kontaktgátlási kísérletekben alkalmazzák. A szokásos tenyésztőközegek gyakran 2–10% szérumot tartalmaznak. A szérum hozzáadása a tenyésztőközeghez a következő célokat szolgálja [21]:
-
A szérum biztosítja a sejtek számára az alapvető tápanyagokat (oldatban és fehérjékhez kötve egyaránt).
-
A szérumban számos, a növekedés elősegítésében és a speciális sejtaktivitásban részt vevő növekedési faktor és hormon található.
-
Számos kötőfehérjét tartalmaz, például albumint és transzferint, amelyek más vegyi anyagokat szállítanak a sejtbe. Az albumin például zsírokat, vitaminokat, hormonokat stb. juttat a sejtekbe.
-
Emellett olyan fehérjéket is biztosít, mint a fibronektin, amelyek fokozzák a sejtek tapadását a szubsztrátumhoz. Ezen felül olyan terjedési elemeket termel, amelyek elősegítik a sejtek osztódás előtti terjeszkedését.
-
Proteázgátlókat juttat a sejtekbe, amelyek megakadályozzák a proteolízist.
-
Emellett olyan ásványi anyagokat is tartalmaz, mint a Na+, K+, Zn2+ és Fe2+.
-
Növeli a táptalaj viszkozitását, így védve a sejteket a szuszpenziós tenyésztés során fellépő keverés okozta mechanikai sérülésektől.
-
Pufferként is működik.
Hivatkozások
[1] Morgan J, Morton H, Parker R. Állati sejtek táplálkozása szövetkultúrában; kezdeti vizsgálatok szintetikus táptalajon. Proc Soc Exp Biol Med. 1950;73:1-8
[2] Kerbel R, Blakeslee D. A magzati borjú szérum egyik komponensének gyors adszorpciója tenyésztett emlőssejtek által. Lehetséges artefaktusforrás a sejtspecifikus antigének elleni antiszérumok vizsgálatában. Immunology. 1976;31:881-91
[3] Sula K, Draber P, Nouza K. A sejtszuszpenziók előállításához használt tápközeghez adott szérum mint lehetséges artefaktusforrás a popliteális nyirokcsomó-teszt segítségével vizsgált sejtmediált reakciókban. J Immunogenet. 1980;7:483-9
[4] Mariani E, Mariani A, Monaco M, Lalli E, Vitale M, Facchini A. Kereskedelmi forgalomban kapható szérummentes táptalajok: hibridómák tenyésztése és monoklonális antitestek előállítása. J Immunol Methods. 1991;145:175-83
[5] Barnes D, Sato G. Módszerek tenyésztett sejtek szérummentes tápközegben történő szaporítására. Anal Biochem. 1980;102:255-70
[6] Yu H, Lu S, Gasior K, Singh D, Vazquez Sanchez S, Tapia O, et al. A HSP70 chaperonok az RNS-mentes TDP-43-at anizotróp, sejtmagon belüli folyékony gömb alakú héjakba kísérik. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, et al. A Down-szindróma által kiváltott szeneszcencia megzavarja az idegsejt-elősejtek sejtmag-szerkezetét. Cell Stem Cell. 2022;29:116–130.e7
[8] Iscove N, Melchers F. A szérum teljes helyettesítése albuminnal, transzferrinnel és szójabab-lipiddel lipopoliszacharidra reagáló B-limfociták tenyészetében. J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. A hibridómák tenyésztésére és monoklonális antitestek előállítására szolgáló, kémiailag definiált, fehérje-mentes tápközeg szisztematikus fejlesztése. J Biotechnol. 1996;45:111-23
[10] Darfler F. Fehérje-mentes táptalaj hibridómák és az immunrendszer egyéb sejtjeinek tenyésztéséhez. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Szérummentes sejtkultúra: egységes megközelítés. Cell. 1980;22:649-55
[12] Hamilton W, Ham R. Kínai hörcsög sejtvonalak klonális szaporodása fehérje-mentes táptalajon. In Vitro. 1977;13:537-47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. A fénynek kitett, HEPES-tartalmú tenyésztőközeg citotoxikus hatásainak elemzése. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Berthois Y, Katzenellenbogen J, Katzenellenbogen B. A szövettenyésztő táptalajokban található fenolvörös gyenge ösztrogén: következmények az ösztrogénre reagáló sejtek tenyészetben történő vizsgálatára nézve. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Szérummentes táptalajok kifejlesztése. In: Master JRW (szerk.). Animal Cell culture, 3. kiadás. Oxford: Oxford University Press; 2000.
[16] Perlman D. Antibiotikumok alkalmazása sejtkultúra-táptalajokban. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. A mikoplazmafertőzés terjedése és ellenőrzése sejtkultúrákban. In Vitro. 1976;12:643-8
[18] Masters J, Stacey G. A tápközeg cseréje és a sejtvonalak átültetése. Nat Protoc. 2007;2:2276–84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M, et al. A KDM6A hiszton-demetiláz közvetlenül érzékeli az oxigént a kromatin és a sejtek sorsának szabályozása érdekében. Science. 2019;363:1217-1222
[20] Molla Kazemiha V, Azari S, Amanzadeh A, Bonakdar S, Shojaei Moghadam M, Habibi Anbouhi M, et al. A Plasmocin™ hatékonysága különböző, mollicutákkal fertőzött emlős sejtvonalakon, összehasonlítva a sejtkultúrában általánosan használt antibiotikumokkal: helyi tapasztalatok. Cytotechnology. 2011;63:609-20
[21] Kragh Hansen U. A ligandum szérumalbuminhoz való kötődésének molekuláris aspektusai. Pharmacol Rev. 1981;33:17-53