
, L-аланил-L-глутаминов разтвор")
Обем: 100 ml
Съхранение: ≤ –15°C
Стерилност: Стерилно филтриран
Стабилен глутаминов разтвор (L-аланил-L-глутамин, 200 mM) е високостабилна дипептидна форма на L-глутамин, предназначена като пряк заместител на конвенционалния L-глутамин в клетъчни културални среди. L-глутаминът е незаменима аминокиселина и основен източник на енергия за култивираните клетки, като играе критична роля в клетъчния растеж, метаболизма и синтеза на протеини.
Приложение и предимства
В стандартните течни среди L-глутаминът се разгражда относително бързо при 37 °C, което води до образуването на токсични странични продукти, като амониеви йони, които могат да повлияят негативно на жизнеспособността на клетките и резултатите от експериментите. Стабилният глутамин преодолява това ограничение, като осигурява неразграждаща се дипептидна форма, която остава непокътната в условия на култивиране.
Клетките ензимно разграждат дипептидната връзка, за да освободят L-глутамин според нуждите, като осигуряват непрекъснато свежо снабдяване и предотвратяват натрупването на вредни отпадни продукти. Това прави разтвора особено подходящ за дългосрочни клетъчни култури и системи за растеж с висока плътност.
Формулировка и употреба
Разтворът L-аланил-L-глутамин се приготвя във вода, подходяща за клетъчни култури, в концентрация 200 mM и се филтрира стерилно за приложения, чувствителни към замърсяване. Може да се разреди директно в пълни среди според експерименталните изисквания. Съхранявайте при ≤ –15 °C и избягвайте повтарящи се цикли на замразяване и размразяване, за да поддържате стабилността на продукта.
Само за изследователски цели. Не е предназначен за употреба в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.
")
Обем: 100 ml
Съхранение: +2°C до +8°C
Стерилност: Стерилно филтриран
HEPES буферен разтвор (1 M), известен също като N-2-хидроксиетилпиперазин-N-2-етансулфонова киселина, е цвиттерионен органичен буферен агент, широко използван в среди за клетъчна култура. Той е предназначен за поддържане на стабилни pH условия във физиологичния диапазон от 6,7 до 8,6, подпомагайки оптималната клетъчна функция по време на ин витро приложения.
Приложение и предимства
HEPES осигурява надеждна буферна способност в системи за клетъчна култура, особено когато клетките се обработват извън CO₂ инкубатор. Добавянето на 10–25 mM HEPES към културалната среда осигурява подобрена pH стабилност по време на продължителни периоди на манипулация, което помага за поддържане на постоянни експериментални условия.
Този буфер е непроницаем за мембрани, има минимална интерференция с биохимичните реакции и демонстрира силна химична и ензимна стабилност. Тези свойства го правят подходящ за широк спектър от клетъчни култури и биохимични приложения.
Формулировка и употреба
Разтворът се доставя като 1 M концентрат, приготвен във вода, подходяща за клетъчни култури, и е стерилно филтриран за употреба в среди, чувствителни към замърсяване. Може да се разреди до желаната работна концентрация в зависимост от изискванията на приложението. Съхранявайте при +2°C до +8°C и боравете в асептични условия, за да запазите целостта на продукта.
Само за изследователски цели. Не е предназначен за употреба в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.
, 250 g/L")
Обем: 50 ml
Съхранение: +2°C до +8°C
Стерилност: Стерилно филтриран
D-(+)-глюкозен разтвор (декстрозен разтвор) е стерилна, готова за употреба добавка, съдържаща естествено срещащата се захар D-(+)-глюкоза, основен компонент на клетъчния метаболизъм. Глюкозата участва в основни биологични процеси като производството на енергия, гликозилирането и образуването на гликани, които допринасят за клетъчната структура и функция.
Приложение и ползи
Този разтвор на глюкоза се използва широко като добавка в среди за клетъчна култура и в многобройни приложения в клетъчната и молекулярната биология. Като основен източник на въглерод и енергия, глюкозата подпомага клетъчния растеж, пролиферацията и метаболитната активност. Участието ѝ в биосинтетичните пътища я прави критично важна за поддържането на нормалната клетъчна физиология и експерименталната последователност.
Състав и употреба
Разтворът се доставя с висока концентрация от 250 g/L глюкоза, което позволява гъвкаво разреждане в културални среди според експерименталните изисквания. Той е стерилно филтриран, за да се гарантира пригодността му за приложения, чувствителни към замърсяване. Съхранявайте при температура от +2°C до +8°C и боравете асептично, за да поддържате качеството и стабилността на продукта.
Само за изследователски цели. Не е предназначен за употреба в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.
, разтвор 100x")
Обем: 10 мл
Съхранение: от +2°C до +8°C
Стерилност: Стерилно филтриран
Разтворът на инсулин-трансферин-селен (ITS) (100x) е химически дефинирана добавка, предназначена за широк спектър от приложения в клетъчните култури. Най-често се използва като добавка към базови среди за клетъчни култури, за да подпомогне клетъчния растеж при условия на намалено или липсващо серум.
Приложение и предимства
Нашата добавка ITS осигурява съставките, необходими за оптималното функциониране на безсерумните среди. Чрез добавяне на ITS към конвенционалните хранителни среди, необходимостта от фетално телешко серум (FBS) за рутинното поддържане на много клетъчни линии може да бъде значително намалена. Това помага за минимизиране на вариациите, свързани с употребата на серум, като същевременно се поддържа постоянен клетъчен растеж и жизнеспособност.
Инсулинът подпомага клетъчното усвояване и метаболизма на ключови хранителни вещества, трансферинът улеснява транспорта на желязо, а селенът допринася за антиоксидантната защита и ензимната активност. Заедно тези компоненти насърчават балансиран клетъчен метаболизъм и подобрена възпроизводимост в дефинирани културални системи.
Състав и употреба
Инсулин-трансферин-селен (ITS) се доставя като 100-кратно концентриран разтвор в балансиран солев разтвор на Earle (EBSS) без фенол червено. За стандартни приложения разредете в съотношение 1:100 в подходящата базова среда, за да постигнете препоръчителната работна концентрация. Съхранявайте при температура от +2 °C до +8 °C и боравете с продукта при асептични условия, за да поддържате стабилността и стерилността му.
Само за изследователски цели. Не се използва за диагностични или терапевтични процедури. Не се използва при хора или животни.
")
Обем: 5 мл
Съхранение: от +2 °C до +8 °C
Стерилност: стерилно филтриран
Разтворът от рекомбинантен човешки инсулин е химически дефинирана добавка, често използвана за култивиране на клетъчни линии от бозайници, включително клетки от яйчници на китайски хамстер (CHO). Този разтвор за клетъчни култури съдържа рекомбинантен човешки инсулин, експресиран в Saccharomyces cerevisiae, което гарантира висока чистота и постоянни резултати при изследователски приложения.
Приложение и предимства
Инсулинът се добавя рутинно към безсерумни и химически дефинирани среди, за да се стимулира клетъчният растеж и продуктивност. Като ключов регулаторен хормон, инсулинът подпомага клетъчното усвояване, използване и съхранение на глюкоза, аминокиселини и мастни киселини. Той също така инхибира разграждането на гликоген, протеини и липиди, като по този начин допринася за подобрена клетъчна жизненост и метаболитна стабилност в културни системи. Химически дефинираната формула подпомага възпроизводимостта и минимизира вариативността в чувствителните работни процеси при клетъчни култури.
Биологични свойства и употреба
Инсулинът е двуверижен полипептиден хормон, естествено произвеждан от β-клетките на панкреатичните островчета. Той има молекулно тегло от приблизително 5800 Da. α
- и β-веригите са свързани чрез две междуверижни дисулфидни връзки, а α-веригата съдържа една вътреверижна дисулфидна връзка. За приложения в клетъчни култури разтворът трябва да се борави при асептични условия и да се съхранява при температура от +2 °C до +8 °C, за да се поддържат стабилността и ефективността.
Само за изследователски цели. Не е предназначен за използване в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.
")
Обем: 100 ml
Съхранение: +2°C до +8°C
Стерилност: Стерилно филтриран
Разтворът на натриев пируват (100 mM) е стерилна, готова за употреба добавка, предназначена да осигури допълнителен, лесно достъпен източник на енергия за клетъчни културални среди. Натриевият пируват играе ключова роля в клетъчния енергиен метаболизъм и подпомага растежа на метаболитно активни и бързо пролифериращи клетки, като например туморни клетки. Добавката може да повиши жизнеспособността на клетките и да спомогне за поддържането на метаболитна стабилност в културалните системи.
Приложение и предимства
Този разтвор се използва широко в рутинната клетъчна култура за обогатяване на средите с пируват и насърчаване на оптимални условия за растеж. Той подпомага производството на АТФ, може да спомогне за намаляване на оксидативния стрес и допринася за подобряване на метаболитната ефективност на култивираните клетки. Произведен във вода, подходяща за клетъчна култура, и стерилно филтриран, продуктът гарантира постоянна качество и възпроизводимост в изследователските работни процеси.
Употреба и съвместимост
Препоръчителната крайна концентрация за повечето приложения за клетъчна култура е 1 mM натриев пируват, постигана чрез разреждане на 100 mM запасен разтвор 1:100 в пълна културална среда. Разтворът е съвместим с широка гама от базови среди и клетъчни линии на бозайници. Съхранявайте при +2°C до +8°C и предпазвайте от замърсяване, за да поддържате стабилността на продукта.
Само за изследователски цели. Не е предназначен за употреба в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.
")
Обем: 100 ml Съхранение: ≤-15°C Стерилност: Стерилно филтриран
Антибиотик/антимикотичен разтвор (100x ) е стерилен, готов за употреба концентрат, предназначен за намаляване на рисковете от микробно замърсяване в клетъчни култури и свързани с тях лабораторни приложения. Този разтвор 100x съдържа утвърдена комбинация от пеницилин, стрептомицин и амфотерицин В
- осигуряваща широк спектър на антимикробна активност срещу грам-положителни и грам-отрицателни бактерии, дрожди и нишковидни гъбички. Формулата е подходяща за използване в еукариотни клетъчни култури, бактериални среди и други чувствителни към замърсяване системи, като подпомага чистите и последователни лабораторни операции.
Приложение и предимства Оптимизиран за рутинни изследователски протоколи, този разтвор се използва широко за поддържане на асептични условия в работните процеси на клетъчни култури. То предлага надеждна работа в чувствителни към замърсяване среди, като помага на изследователите да намалят риска от свръхрастеж на микроорганизми, без да се компрометира здравето на клетките или възпроизводимостта на експериментите. Стерилно-филтрираният състав елиминира необходимостта от допълнителни етапи на разтваряне, като подпомага рационализираното приготвяне на среди и намалява променливостта в ежедневните лабораторни процедури.
Употреба и съвместимост За да постигнете стандартни работни концентрации, разредете разтвора в съотношение 1:100 в пълна хранителна среда. Продуктът е съвместим с широк спектър от клетъчни линии на бозайници и базови среди. С постоянната наличност на склад изследователите се възползват от надеждна непрекъснатост на доставките и опростено логистично планиране. За да се запази стабилността, разтворът трябва да се съхранява при ≤ -15 °C и да се предпазва от многократни цикли на замразяване и размразяване. Само за изследователска употреба. Не е предназначен за използване в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.
")
Обем: 100 ml Съхранение: +Стерилност: 2°C до +8°C Стерилно филтриран
MEM Non-Essential Amino Acids (100x ) е стерилна добавка, предназначена да подобри клетъчния растеж и жизнеспособността в системи за клетъчни култури от бозайници. Формулата съответства на 100-кратен концентрат на неесенциалните аминокиселини, открити в стандартната минимална есенциална среда (MEM), което позволява директно добавяне на базови среди с минимална подготовка.
Приложение и предимства Тази добавка осигурява допълнителен аминокиселинен пул за бързо размножаващи се клетки или за клетъчни линии, които са загубили способността си да синтезират несъществени аминокиселини de novo. Чрез облекчаване на метаболитната тежест на биосинтезата тя подпомага подобрената кинетика на растежа, удължената жизнеспособност и по-голямата експериментална последователност
- особено при чувствителни към хранителни вещества култури или култури с висока плътност.
Състав и употреба Разтворът включва глицин, L-аланин, L-аспарагин, L-аспарагинова киселина, L-глутаминова киселина, L-пролин и L-серин. Съвместим е с MEM и повечето други стандартни среди. За да използвате, разредете 1:100 в крайната хранителна среда. Този продукт е стерилно филтриран и готов за употреба без допълнителни стъпки за обработка. Само за изследователска употреба. Не е предназначен за използване в диагностични или терапевтични процедури. Не е предназначен за употреба при хора или животни.

Accutase е готов за употреба, стерилно филтриран разтвор за отделяне на клетки, създаден като щадяща алтернатива на трипсин/EDTA за отделяне на адхезивни клетки от бозайници от стандартни пластмасови съдове за тъканни култури и повърхности с адхезивно покритие. Той съчетава протеолитична и колагенолитична ензимна активност в балансиран солен разтвор, за да осигури ефективно, но контролирано отделяне, като запазва протеините на клетъчната повърхност и подпомага висока жизнеспособност след пасаж и бързо повторно прикрепване.
Формулата на Accutase се основава на фосфатно-буфериран физиологичен разтвор на Дълбеко (DPBS) с EDTA и фенол червено като визуален индикатор за pH. Ензимите са от небозайнически и небактериален произход, което прави Accutase особено подходящ за изследвания на стволови клетки, работни процеси при ваксини и всякакви приложения, при които трябва да се сведе до минимум наличието на замърсители от животински или микробен произход. Разтворът се самоинхибира при 37 °C, така че след отделянето не се изисква неутрализиращ реагент или среда, съдържаща серум – клетките могат да се прехвърлят директно в свежа среда.
Основни характеристики
Готова за употреба 1x стерилно филтрирана течност – не се изисква разреждане или възстановяване
Комбинирана протеолитична и колагенолитична ензимна активност за нежно разграждане
Всяка партида е стандартизирана до определена дисоциационна активност за последователност между партидите
Ензими от небозайнически и небактериален произход
Автоинхибира се при 37 °C – не се изисква неутрализиращ разтвор
Формулиран в PBS на Дълбеко с EDTA
Включен фенол червен като визуален индикатор за pH
pH 6,8 – 7,8
Типични приложения
Accutase нежно дисоциира широка гама от адхезивни и чувствителни клетъчни типове, включително човешки ембрионални стволови клетки (hESCs), човешки индуцирани плюрипотентни стволови клетки (iPSCs), невронални стволови клетки, първични неврони и рутинно култивирани адхезивни линии като HeLa, HEK 293, CHO, MDCK, Vero, NIH/3T3, BHK-21 и A549. Типични примери за употреба включват:
Рутинно субкултивиране и пасажиране на адхезивни клетки от бозайници
Нежно разграждане на единични клетки от hESCs, iPSCs и други чувствителни линии
Подготовка на проби за проточна цитометрия и FACS анализ
Анализ на маркери на клетъчната повърхност, при които целостта на епитопа е от значение
Тестове за клетъчна миграция, пролиферация и апоптоза
Тестове за покой чрез лишаване от серум и проучвания за трансфекция на онкогени
Тестове за миграция на туморни клетки и клетки от невралния гребен
Увеличаване на производството в биореакторни работни процеси
За рутинна работа нанесете приблизително 10 ml Accutase на 75 cm2 културална повърхност и инкубирайте в продължение на 5–10 минути при стайна температура. Оптималното време за инкубиране трябва да се определи за всяка клетъчна линия и не трябва да надвишава един час. Преди добавянето изплакнете клетъчния слой с разтвор на соли без Ca2+/Mg2+, като например DPBS без калций и магнезий, за да отстраните остатъчното серум и двувалентните катиони.
Работа и съхранение
Съхранявайте затворената бутилка замразена при -15 °C или по-ниска температура. Размразявайте при стайна температура или през нощта при +2 °C до +8 °C. Не размразявайте Accutase във водна баня при 37 °C, тъй като повишените температури намаляват ензимната активност. След размразяване разтворът може да се съхранява до 2 месеца при +2 °C до +8 °C; не съхранявайте при стайна температура. Не загрявайте реагента до 37 °C преди употреба – добавете го директно към измитите клетки при стайна температура. За дълъг срок на съхранение се препоръчва разделяне на аликвоти за еднократна употреба, за да се избегнат повтарящи се цикли на размразяване. Винаги работете при асептични условия.
Качество
Произведен съгласно строги стандарти за качество. Всяка партида Accutase е стерилно филтрирана и тествана за стерилност, pH, външен вид и дисоциационна активност, за да се гарантира постоянна и възпроизводима ефективност от партида до партида.
Спецификации на продукта
Спецификация
Подробности
Тип на продуктаРеагент за отделяне/дисоциация на клетки
ФорматСтерилно филтрирана течност, готова за употреба
Обем100 мл
Работна концентрация1x (готова за употреба)
Ензимна активностКомбинирана протеолитична и колагенолитична
Произход на ензимаНе от бозайници и не от бактерии
Буферна системаPBS на Дълбеко с EDTA
pH индикаторФенол червено
pH диапазон6,8 – 7,8
Външен видПрозрачен разтвор с цвят от бледочервен до оранжев
Температура на съхранение-15 °C или по-ниска
Стабилност след размразяванеДо 2 месеца при +2 °C до +8 °C
Препоръчително количество за употреба~10 мл на 75 cm2 културална повърхност
Типично време за инкубация5 – 10 минути при стайна температура
Условия за транспортиранеЗамразено в сух лед
ПредназначениеСамо за научноизследователски цели и по-нататъшно производство
Формулировка (състав на литър)
Компонент
Концентрация (mg/L)
Неорганични соли
Натриев хлорид (NaCl)8000,00
Динатриев хидрогенфосфат (Na2HPO4)1150,00
Калиев хлорид (KCl)200,00
Калиев дихидрогенфосфат (KH2PO4)200,00
Други компоненти
EDTA · 4Na (тетранатриев EDTA)220,00
Фенол червено3,00
Патентована ензимна смес (протеолитична и колагенолитична активност)1x
Accutase е регистрирана търговска марка на Innovative Cell Technologies, Inc.
, w: 2 mM L-Glutamine, w: 2,2 g/L NaHCO3, w: EBSS")
Тази готова за употреба, стерилно филтрирана течна формула е допълнена с балансиран солен разтвор на Ърл (EBSS), 2 mM L-глутамин, D-глюкоза (1,0 g/L) и 2,2 g/L натриев бикарбонат (NaHCO3), което я прави подходяща за употреба в инкубаторна атмосфера с контролирано съдържание на CO2 (обикновено 5 % CO2). Включеният фенол червен действа като индикатор за pH, което позволява удобно визуално наблюдение на състоянието на средата по време на клетъчната култура.
Основни характеристики
Класическа формула на MEM на Игъл с балансиран солен разтвор на Ърл (EBSS)
Включен 2 mM L-глутамин – готов за незабавна употреба
2,2 g/L натриев бикарбонат – буфериран за инкубация при 5 % CO2
С D-глюкоза (1,0 g/L) като основен източник на въглерод
С фенол червено като индикатор за pH
Без HEPES и без натриев пируват
Стерилно филтрирана течна среда, готова за употреба
pH 7,0 – 7,6
Типични приложения
EMEM поддържа култивирането на голямо разнообразие от клетъчни линии на бозайници, включително HeLa, HEK 293, Vero, MRC-5, L-929, BHK-21 и много първични клетки. Общите приложения включват:
Рутинна поддръжка и разширяване на адхезивни клетъчни линии
Работни процеси за размножаване на вируси и производство на ваксини
Приложения за цитотоксичност и биологични тестове
Изследвания на трансфекция и протеинова експресия
Фундаментални изследвания в клетъчната и молекулярната биология
За оптимален клетъчен растеж, EMEM обикновено се допълва с 5–10 % фетално телешко серум (FBS) и, в зависимост от клетъчната линия, с неesenциални аминокиселини (NEAA) и антибиотици като пеницилин/стрептомицин.
Работна среда и съхранение
Съхранявайте затворената бутилка при температура от +2 °C до +8 °C, защитена от светлина. След отваряне използвайте при асептични условия. L-глутаминът в разтвора подлежи на постепенно разграждане – за най-добри резултати препоръчваме да използвате средата в рамките на 4 седмици след отваряне или да я допълните с пресен L-глутамин преди употреба, ако се съхранява за по-дълъг период. Оставете средата да се затопли до 37 °C, преди да я добавите към клетките.
Качество
Произведено съгласно строги стандарти за качество. Всяка партида се тества за стерилност, pH, осмолалност и нива на ендотоксини, за да се гарантира постоянна ефективност при приложения в клетъчни култури.
Спецификации на продукта
Спецификация
Подробности
Тип на продуктаMEM
Категория на продуктаСреди за клетъчна култура
ФорматТечност
СтерилнаДа
Размер500 мл
L-глутаминС L-глутамин (2 mM)
ГлюкозаС глюкоза (1,0 g/L)
Натриев бикарбонатС NaHCO3 (2,2 g/L)
HEPESБез HEPES
Натриев пируватБез натриев пируват
Фенол червеноС фенол червено
Солев разтворБалансиран солен разтвор на Ърл (EBSS)
pH7,0 – 7,6
Съдържание на ендотоксиниНе е посочено
СъхранениеОт +2 °C до +8 °C
Формулировка (състав на литър)
Компонент
Концентрация (mg/L)
Неорганични соли
Калциев хлорид · 2H2O265,00
Магнезиев сулфат97,72
Калиев хлорид400,00
Натриев хлорид6 800,00
Натриев дихидрогенфосфат, безводен122,00
Натриев бикарбонат (NaHCO3)2 200,00
Аминокиселини
L-аргинин · HCl126,00
L-цистин · 2HCl31,30
L-глутамин292,00
L-хистидин · HCl · H2O42,00
L-изолевцин52,00
L-лейцин52,00
L-лизин · HCl72,50
L-метионин15,00
L-фенилаланин32,00
L-треонин48,00
L-триптофан10,00
L-тирозин · 2Na · 2H2O51,90
L-валин46,00
Витамини
D-калциев пантотенат1,00
Холин хлорид1,00
Фолиева киселина1,00
мио-инозитол2,00
Никотинамид1,00
Пиридоксал · HCl1,00
Рибофлавин0,10
Тиамин · HCl1,00
Други компоненти
D(+)-глюкоза1 000,00
Фенол червено10,00

Основните характеристики на Freeze Medium CM-1 включват:
Широка съвместимост: Ефективна за широк спектър от клетъчни типове, включително първични клетки, стволови клетки и установени клетъчни линии.
Висока жизнеспособност: Оптимизирана за максимално възстановяване на клетките след размразяването им и тяхната жизнеспособност, което гарантира надеждни експериментални резултати.
Готов за употреба: Удобно подготвени и стерилизирани за незабавно приложение, което намалява времето за подготовка и риска от замърсяване.
Повишена стабилност: Поддържа постоянна производителност при стандартни условия на криоконсервация, като осигурява възпроизводими резултати.
Дълъг срок на съхранение: CM-1 е готова за употреба среда за криоконсервация, съдържаща серум, която може да се съхранява в хладилник до една година.
Използване на CM-1 за замразяване на клетки
За да използвате CM-1 за замразяване на адхезивни и суспензионни клетки, следвайте следните стъпки
За адхерентни клетки ги измийте и ги отделете от субстрата за култивиране. За суспензионни клетки преминете директно към следващата стъпка.
Пребройте клетките, за да се уверите, че те са с подходяща концентрация.
Центрофугирайте клетките, за да ги пелетирате, след което ги ресуспендирайте в среда за замразяване CM-1.
Прехвърлете ресуспендираните клетки в криовиали.
Използвайте метод за бавно замразяване, преди да прехвърлите клетките за дългосрочно съхранение
Метод
Описание
Стъпки
❄️
Ръчно замразяване
Метод, включващ постепенно намаляване на температурата, за да се осигури жизнеспособността на клетките
1️⃣ Поставете клетките в среда за замразяване във фризер с температура 4 °C за 40 минути.
2️⃣ Прехвърлете във фризер с температура -80°C за 24 часа.
3️⃣ Съхранявайте клетките в течен азот за дългосрочно запазване
❄️
Използване на Mr. Frosty
Удобно устройство, което позволява контролирана скорост на замразяване без електрическо захранване
1️⃣ Подгответе клетките в криовиали със замразяваща среда.
2️⃣ Поставете криовиолите в контейнера на Mr. Frosty.
3️⃣ Съхранявайте при -80 °C за 24 часа, преди да прехвърлите в течен азот
❄️
Фризер с контролирана скорост
Високопрецизен фризер на Thermo Fisher или на други производители, предназначен за контролирано намаляване на температурата
1️⃣ Програмирайте устройството за постепенно намаляване на температурата.
2️⃣ Поставете подготвените клетки във фризера.
3️⃣ След цикъла на замразяване прехвърлете клетките в течен азот
Съхранявайте криовиолите при температури под -130 °C или в течен азот за дългосрочно съхранение.
Съставки
Съдържа FBS, DMSO, глюкоза, соли
Буфериращ капацитет: pH = 7,2 до 7,6
Средата за замразяване CM-1 на Cytion предлага надеждно решение за криоконсервация, като осигурява висока клетъчна жизнеспособност и функционалност след размразяване за широк спектър от изследователски приложения.

Средата Ham's F-12K (Kaighn's) е внимателно формулирана, за да оптимизира условията за клетъчно култивиране. Тя се отличава с обогатен състав, осигуряващ повишени нива на основни компоненти като аминокиселини и натриев пируват, както и допълнителни елементи, включително путресцин, тимидин, хипоксантин и цинк. Тези добавки дават възможност на изследователите да допълват средата с минимално количество серум или определени компоненти за специфични клетъчни типове, което улеснява прецизните експериментални условия.
Забележително е, че средата Ham's F-12K (Kaighn's) не съдържа протеини или растежни фактори. Следователно често е необходимо добавяне на растежни фактори и фетален говежди серум (FBS), което позволява на изследователите да адаптират средата към изискванията на техните специфични клетъчни линии. За оптимална ефективност концентрацията на FBS трябва да бъде внимателно оптимизирана за всяка клетъчна линия, като се гарантира оптимален растеж и функционалност.
За поддържане на физиологично рН в средата Ham's F-12K (Kaighn's) се използва буферна система от натриев бикарбонат (2,5 g/L), което налага контролирана среда от 5-10 % CO2 по време на култивирането. Това гарантира, че рН на средата остава в идеалния диапазон за клетъчен растеж и жизнеспособност
Контрол на качеството
pH = 7,2 +/
- 0,02 при 20-25°C.
Всяка партида е тествана за стерилност и липса на микоплазма и бактерии.
Поддръжка
Съхранявайте в хладилник на тъмно при температура от +2°C до +8°C. Замразяването, както и затоплянето до +37°C намаляват до минимум качеството на продукта.
Не нагрявайте средата до повече от 37°C и не използвайте неконтролируеми източници на топлина (напр. микровълнови уреди).
Ако трябва да се използва само част от средата, извадете това количество от бутилката и го загрейте на стайна температура.
Срокът на годност на всяка среда, с изключение на основната среда, е 8 седмици от датата на производство.
Състав
Компоненти
mg/L
Неорганични соли
Калциев хлорид x 2H2O
135,24
Меден(II) сулфат x 5H2O
0,00
Железен (II) сулфат x 7H2O
0,83
Магнезиев хлорид x 6H2O
105,72
Магнезиев сулфат x 7H2O
394,49
Калиев хлорид
283,29
Калиев дихидрогенфосфат
58,52
Натриев хлорид
7597,20
натриев хидрогенфосфатбезводен
115,02
Цинков сулфат x 7H2O
0,14
Други компоненти
D(+)-глюкоза, безводна
1260,00
Хипоксантин
4,08
DL-α-липоева киселина
0,21
Червен фенол
3,00
Путресцин x 2HCl
0,32
Натриев пируват
220,00
NaHCO3
2500,00
Тимидин
0,73
Аминокиселини
L-аланин
17,82
L-аргинин х HCl
421,40
L-аспарагин x H2O
30,02
L-аспарагинова киселина
26,62
L-цистеин x HCl x H2O
70,24
L-глутамин
292,20
L-глутаминова киселина
29,42
Глицин
15,01
L-хистидин х HCl х H2O
41,92
L-изолевцин
7,87
L-Leucine
26,24
L-лизин x HCl
73,04
L-метионин
8,95
L-фенилаланин
9,91
L-пролин
69,06
L-серин
21,02
L-треонин
23,82
L-триптофан
4,08
L-тирозин
10,87
L-Valine
23,42
Витамини
D(+)-Биотин
0,07
D-калциев пантотенат
0,48
Холин хлорид
13,96
Фолиева киселина
1,32
мио-инозитол
18,02
Никотинамид
0,04
Пиридоксин x HCl
0,06
Рибофлавин
0,04
Тиамин x HCl
0,34
Витамин В12
1,36

Фосфатно-буферираният физиологичен разтвор (PBS) е широко използван буферен разтвор в биологичните и химичните изследвания. Той играе решаваща роля за поддържане на pH баланса и осмоларността по време на различни експериментални процедури, включително обработка на тъкани и клетъчни култури. Нашият разтвор на PBS е щателно формулиран с висококачествени съставки, за да се гарантира стабилност и надеждност при всеки експеримент. Осмоларността и концентрацията на йони в нашия разтвор PBS са близки до тези в човешкото тяло, което го прави изотоничен и нетоксичен за повечето клетки.
Състав на нашия разтвор на PBS
Нашият разтвор на PBS е смес от свръхчисти фосфатни буфери и физиологични разтвори с коригирано pH. В работна концентрация 1Х той съдържа:
8000 mg/L Натриев хлорид (NaCl)
200 mg/L Калиев хлорид (KCl)
1150 mg/L Натриев фосфат двуосновен безводен (Na2HPO4)
200 mg/L Калиев фосфат монобазичен безводен (KH2PO4)
Този състав осигурява оптимално рН и йонно равновесие, подходящи за широк спектър от биологични приложения.
Приложения на нашия разтвор на PBS
Нашият разтвор на PBS е идеален за различни приложения в биологичните изследвания. Неговите изотонични и нетоксични свойства го правят подходящ за разреждане на вещества и за изплакване на клетъчни контейнери. Разтворите PBS, съдържащи EDTA, са ефективни за отделяне на прикрепени и слепени клетки. Въпреки това към PBS не трябва да се добавят двувалентни метали, като например цинк, тъй като това може да доведе до утаяване. В такива случаи се препоръчват буферите на Good's. Освен това нашият разтвор на PBS е приемлива алтернатива на вирусната транспортна среда за транспортиране и съхранение на РНК вируси, включително SARS-CoV-2.
Контрол на качеството
Стерилно филтриран
Съхранение и срок на годност
Съхранявайте при температура от +2°C до +25°C, защитени от светлина.
След отваряне съхранявайте при 2°C до 25°C и използвайте в рамките на 24 месеца.
Условия за транспортиране
Температура на околната среда
Поддръжка
Съхранявайте в хладилник при температура от +2°C до +8°C на тъмно. Избягвайте замразяването и честото затопляне до +37°C, тъй като това намалява качеството на продукта.
Не нагрявайте средата над 37°C и не използвайте неконтролирани източници на топлина, като например микровълнови уреди.
Ако трябва да се използва само част от средата, отстранете необходимото количество и го загрейте до стайна температура преди употреба.
Състав
Категория
Компоненти
Концентрация (mg/L)
Соли
Калиев хлорид
200
Калиев фосфат монобазичен безводен
200
Натриев хлорид
8000
Натриев фосфат двуосновен безводен
1150

Първоначално създадена за поддържане на растежа на човешки левкемични клетки в суспензионни и монослойни култури, средата RPMI 1640 еволюира чрез модификации от изследователи и търговски доставчици, за да стане подходяща за разнообразни клетки на бозайници. Тя е изключително съвместима с клетъчни линии като HeLa, Jurkat, MCF-7, PC12, PBMC, астроцити и карциноми.
Средата RPMI 1640 се отличава от другите среди за клетъчни култури поради уникалния си състав. Тя съдържа значително количество фосфати, аминокиселини и витамини. По-специално, тя включва биотин, витамин В12 и PABA, които отсъстват в Eagle's Minimal Essential Medium или Dulbecco's Modified Eagle Medium. Освен това средата RPMI 1640 показва значително повишени концентрации на витамините инозитол и холин. Тя обаче не съдържа протеини, липиди или растежни фактори. Поради това обикновено се налага добавяне на 10% фетален говежди серум (FBS), за да се осигурят оптимални условия за клетъчен растеж.
Буферната система на средата RPMI 1640 разчита на натриев бикарбонат и изисква 5-10% CO2 среда, за да се поддържа физиологично подходящо pH. Включването на редуциращия агент глутатион допълнително отличава тази среда от останалите.
Контрол на качеството
Стерилно филтрирани
Съхранение и срок на годност
Съхранявайте при температура от +2°C до +8°C, защитена от светлина.
След отваряне съхранявайте при 4°C и използвайте в рамките на 6-8 седмици.
Условия за транспортиране
Температура на околната среда
Поддръжка
Съхранявайте в хладилник при температура от +2°C до +8°C на тъмно. Избягвайте замразяването и честото затопляне до +37°C, тъй като това намалява качеството на продукта.
Не нагрявайте средата над 37°C и не използвайте неконтролирани източници на топлина, като например микровълнови уреди.
Ако трябва да се използва само част от средата, отстранете необходимото количество и го загрейте до стайна температура преди употреба.
Състав
Категория
Компоненти
Концентрация (mg/L)
Аминокиселини
Глицин
10.00
L-аланил-L-глутамин
434.40
L-аргинин
200.00
L-аспарагинH2O
56.82
L-аспарагинова киселина
20.00
L-цистин 2HCl
65.20
L-глутаминова киселина
20.00
L-Histidine HClH2O
20.27
L-хидрокси-L-пролин
20.00
L-изолевцин
50.00
L-Leucine
50.00
L-лизин HCl
40.00
L-метионин
15.00
L-фенилаланин
15.00
L-пролин
20.00
L-серин
30.00
L-Threonine
20.00
L-триптофан
5.00
L-тирозин 2Na 2H2O
28.83
L-Valine
20.00
Витамини
р-аминобензоена киселина
1.00
D-биотин
0.20
Холин хлорид
3.00
D-калциев пантотенат
0.25
Фолиева киселина
1.00
мио-Инозитол
35.00
Никотинамид
1.00
Пиридоксин HCl
1.00
Рибофлавин
0.20
Тиамин HCl
1.00
Витамин В12
0.005
Неорганични соли
Ca(NO3)2 4H2O
100.00
KCl
400.00
MgSO4 7H2O
100.00
NaCl
6000.00
NaHCO3
2000.00
Na2HPO4
800.00
Други компоненти
D-глюкоза
2000.00
L-глутатион, редуциран
1.00
Фенол червено натриева сол
5.30
, w: 3,1 g/L глюкоза, w: 2,5 mM L-глутамин, w: 15 mM HEPES, w: 0,5 mM натриев пируват, w: 1,2 g/L NaHCO3")
Тази уникална формула комбинира Dulbecco's Modified Eagle Medium (DMEM) и Ham's F-12 (Ham's Nutrient Mixture F-12) в точно съотношение 1:1. Добавянето на L-глутамин допълнително подобрява състава ѝ.
DMEM, произхождаща от минималната есенциална среда на Игъл (EMEM), предлага повишена концентрация на аминокиселини и витамини в сравнение с предшественика си. За разлика от нея, Ham's F-12 се базира на средата Ham's F-10, като осигурява допълващ набор от есенциални компоненти.
За да се подпомогне оптималният клетъчен растеж, обичайна практика е DMEM:Ham's F12 да се допълва с FBS при типична концентрация от 5-10%. Това добавяне е необходимо, тъй като средата не съдържа растежни хормони, липиди и протеини, които са от решаващо значение за клетъчното развитие.
DMEM:Ham's F12 включва pH буферна система и често се допълва с фенол червено, индикатор за pH. Култивираните клетки в DMEM:Ham's F12 или всяка друга среда, използваща бикарбонатна буферна система, изискват контролирана CO2 среда от 5-10%, за да се поддържат подходящи нива на pH.
Контрол на качеството
Стерилно филтрирана
Съхранение и срок на годност
Съхранявайте при температура от +2°C до +8°C, защитено от светлина.
След отваряне съхранявайте при 4°C и използвайте в рамките на 6–8 седмици.
Условия за транспортиране
Околна температура
Съхранение
Съхранявайте в хладилник при температура от +2°C до +8°C на тъмно. Избягвайте замразяване и често затопляне до +37°C, тъй като това намалява качеството на продукта.
Не загрявайте средата над 37 °C и не използвайте неконтролирани източници на топлина, като например микровълнови уреди.
Ако ще се използва само част от средата, извадете необходимото количество и го затоплете до стайна температура преди употреба.
Състав
Категория
Компоненти
Концентрация (mg/L)
Аминокиселини
Глицин
18,75
L-аланин
4,45
L-аргинин HCl
147,50
L-аспарагин H₂O
7,50
L-аспарагинова киселина
6,65
L-цистеин HCl H₂O
17,56
L-цистин 2 HCl
31,29
L-глутаминова киселина
7,35
L-глутамин
365,00
L-хистидин HCl H₂O
31,48
L-изолевцин
54,47
L-лейцин
59,05
L-лизин HCl
91,25
L-метионин
17,24
L-фенилаланин
35,48
L-пролин
17,25
L-серин
26,25
L-треонин
53,45
L-триптофан
9,02
L-тирозин 2 Na 2 H2O
55,79
L-валин
52,85
Витамини
D-биотин
0,0035
Холин хлорид
8,98
D-калциев пантотенат
2,24
Фолиева киселина
2,66
мио-инозитол
12,60
Никотинамид
2,02
Пиридоксин HCl
0,031
Пиридоксал HCl
2,00
Рибофлавин
0,219
Тиамин HCl
2,17
Витамин В12
0,68
Неорганични соли
CaCl2 2 H2O
154,50
CuSO4 5 H2O
0,0013
Fe(NO3)3 9 H2O
0,05
FeSO4 7 H2O
0,417
KCl
311,80
MgCl2 6 H2O
61,20
MgSO4 7 H2O
100,00
NaCl
6996,00
NaHCO3
1200,00
Na2HPO4
71,02
NaH2PO4 2 H2O
70,87
ZnSO4 7 H2O
0,432
Други компоненти
D-глюкоза
3151,00
Хипоксантин
2,40
HEPES
3574,50
Линоленова киселина
0,042
Липоева киселина
0,105
Натриева сол на фенол червено
8,63
Путресцин 2 HCl
0,081
Натриев пируват
55,00
Тимидин
0,365
, w: 3,0 g/L глюкоза, w: стабилен глутамин, w: 2,0 mM натриев пируват, w: 2,2 g/L NaHCO3")
- 0,02 при 20-25 °C. Всяка партида е тествана за стерилност и отсъствие на микоплазма и бактерии. Съхранение Съхранявайте в хладилник при температура от +2°C до +8°C на тъмно място. Замразяването и затоплянето до +37°C намаляват качеството на продукта. Не загрявайте средата до повече от 37°C и не използвайте неконтролируеми източници на топлина (например микровълнови уреди). Ако ще се използва само част от средата, извадете това количество от бутилката и го затоплете до стайна температура. Срокът на годност за всички среди, с изключение на основната среда, е 6 до 8 седмици от датата на отваряне. Състав Съставки mg/L Неорганични солиКалциев хлорид x 2H2O132,00 Магнезиев сулфат97,67 Калиев хлорид400 Натриев хлорид6460 Динатриев хидроген фосфат (безводен)504 Други компонентиD(+)-глюкоза (безводен)3 000 Глутатион (редуциран)0 Месен пептон600 Фенол червена натриева сол11 АминокиселиниL-аланин13 L-аргинин x HCl42,14 L-аспарагин x H2O45,03 L-аспарагинова киселина19,97 L-цистеин x HCl x H2O31,75 L-глутамин (стабилен)219,15 L-глутаминова киселина22 Глицин7 L-хистидин x HCl x H2O20,96 L-хидроксипролин19,67 L-изолевцин39,3 L-левцин39,36 L-лизин x HCl36,54 L-метионин14,92 L-фенилаланин16,52 L-пролин17,27 L-серин26,28 L-треонин17,87 L-триптофан3,06 L-тирозин динатриева сол26,10 L-валин17,57 Витаминиp-Аминобензоена киселина1 Аскорбинова киселина0,56 D(+)-Биотин0,20 D-калциев пантотенат0,20 Холин хлорид5 Фолиева киселина10 Мио-инозитол36 Никотинамид0 Никотинова киселина0,5 Пиридоксал HCl0,5 Пиридоксин HCl0,50 Рибофлавин0,20 Тиамин HCl0 Витамин В122

Среда 199 предлага редица приложения в областта. Тя може ефективно да поддържа комплекса кумулус-оцит (COC) и да подпомага ин витро съзряването на яйцеклетки. Освен това тя се използва при промиване на аспирационни линии по време на събиране на яйцеклетки от немски крави от породата Холщайн. Освен това среда 199 служи като отлична среда за култивиране на сърдечни ендотелни клетки, получени от плъхове. Тези приложения демонстрират гъвкавостта и приспособимостта на среда 199 към различни експериментални нужди.
История
Разработването на среда 199 през 50-те години на миналия век бележи значителен напредък в областта на средите за тъканни култури. Преди нейното въвеждане много среди за култивиране са разчитали на продукти от животински произход и тъканни екстракти. Морган и колегите му обаче правят революция в тази област, като формулират напълно определен хранителен източник за клетъчни култури. Чрез своите експерименти, включващи различни комбинации от витамини, аминокиселини и други фактори, те откриват изключителните свойства на среда 199, стимулираща растежа.
Контрол на качеството
pH = 7,2 +/
- 0,02 при 20-25°C.
Всяка партида е тествана за стерилност и липса на микоплазма и бактерии.
Поддръжка
Съхранявайте в хладилник на тъмно при температура от +2°C до +8°C. Замразяването и затоплянето до +37°С намаляват до минимум качеството на продукта.
Не нагрявайте средата до повече от 37°С и не използвайте неконтролируеми източници на топлина (напр. микровълнови уреди).
Ако трябва да се използва само част от средата, извадете това количество от бутилката и го загрейте на стайна температура.
Срокът на годност на всяка среда, с изключение на основната среда, е 8 седмици от датата на производство.
Състав
Компоненти
mg/L
Неорганични соли
Калциев хлорид x 2H2O
264,92
Железен (III) нитрат x 9H2O
0,72
Магнезиев сулфат
97,67
Калиев хлорид
400,00
Натриев ацетат x 3H2O
82,95
Натриев хлорид
6,800.00
Натриев дихидрогенфосфат x H2O
140,00
Други компоненти
Аденин сулфат
10,00
AMP
0,20
ATP
1,00
Холестерол
0,20
2'-дезоксирибоза
0,50
D(+)-глюкоза, безводна
1,000.00
Глутатион (червен)
0,05
Гуанин х HCl
0,30
Хипоксантин
0,30
Фенол червено
10,00
D-рибоза
0,50
Тимин
0,30
Tween 80
4,90
Урацил
0,30
Ксантин
0,30
NaHCO3
2,200.00
Аминокиселини
L-аланин
25,00
L-аргинин x HCl
70,00
L-аспарагинова киселина
30,00
L-цистеин x HCl x H2O
0,10
L-цистин
20,00
Стабилен L-глутамин
149,00
L-глутаминова киселина
67,00
Глицин
50,00
L-хистидин x HCl x H2O
21,88
L-хидроксипролин
10,00
L-изолевцин
20,00
L-Leucine
60,00
L-лизин x HCl
70,00
L-метионин
15,00
L-фенилаланин
25,00
L-пролин
40,00
L-серин
25,00
L-треонин
30,00
L-триптофан
10,00
L-тирозин
40,00
L-валин
25,00
Витамини
4-аминобензоена киселина
0,05
Аскорбинова киселина
0,05
D(+)-Биотин
0,01
Калциферол
0,10
D-калциев пантотенат
0,01
Холин хлорид
0,50
Фолиева киселина
0,01
мио-Инозитол
0,05
Менадион
0,01
Никотинова киселина
0.025
Никотинамид
0.025
Пиридоксал х HCl
0.025
Пиридоксол х HCl
0.025
Рибофлавин
0,01
DL-α-токоферол фосфат динатриева сол
0,01
Тиамин х HCl
0,01
Витамин А ацетат
0,14

IMDM е подходящ за бързо пролифериращи клетъчни култури с висока плътност, включително Jurkat, COS-7 и макрофаги. Различните модификации на IMDM, налични за редица приложения за клетъчни култури, могат да бъдат намерени с помощта на инструмента за избор на среди. Течните среди осигуряват основни хранителни вещества за всички приложения за клетъчни култури. Всяка от нашите висококачествени среди за клетъчни култури се произвежда в съответствие с първоначално публикуваната формула или модификации, необходими за постоянното функциониране и стабилност на средата за култура.
IMDM срещу DMEM
IMDM съдържа калиев нитрат вместо железен нитрат и HEPES и натриев пируват. Допълнителните компоненти в IMDM я правят по-подходяща за специализирани клетъчни типове и специфични приложения, отколкото DMEM.
IMDM срещу RPMI
IMDM и RPMI имат различни формули, които могат да бъдат от значение за стимулирането с PMA/йономицин. Една от съществените разлики е концентрацията на Ca2+. Докато RPMI съдържа 0,42 mM Ca2+, IMDM съдържа 1,49 mM.
Контрол на качеството
pH = 7,2 +/
- 0,02 при 20-25°C.
Всяка партида е тествана за стерилност и липса на микоплазма и бактерии.
Поддръжка
Съхранявайте в хладилник на тъмно при температура от +2°C до +8°C. Замразяването и затоплянето до +37°С намаляват до минимум качеството на продукта.
Не нагрявайте средата до повече от 37°С и не използвайте неконтролируеми източници на топлина (напр. микровълнови уреди).
Ако трябва да се използва само част от средата, извадете това количество от бутилката и го загрейте на стайна температура.
Срокът на годност на всяка среда, с изключение на основната среда, е 8 седмици от датата на производство.
Състав
Компоненти
mg/L
Неорганични соли
Калциев хлорид x 2 H2O
219,00
Калиев хлорид
330,00
Калиев нитрат
0.076
Магнезиев сулфат, безводен
97,73
Натриев хлорид
4,505.00
Натриев дихидрогенфосфат, безводен
109,00
Натриев селенит
0,02
Други компоненти
D(+)-глюкоза, безводна
4,500.00
HEPES
5,958.00
Натриев пируват
110,00
Фенол червено
15,00
Аминокиселини
L-аланин
25,00
L-аргинин x HCl
84,00
L-аспарагин x H2O
25,00
L-аспарагинова киселина
30,00
L-цистин х 2HCl
91,24
L-глутамин
584,00
L-глутаминова киселина
75,00
Глицин
30,00
L-хистидин x HCl x H2O
42,00
L-изолевцин
104,80
L-Leucine
104,80
L-лизин x HCl
146,20
L-метионин
30,00
L-фенилаланин
66,00
L-пролин
40,00
L-серин
42,00
L-треонин
95,20
L-триптофан
16,00
L-тирозин x 2Na
104,20
L-Valine
93,60
Витамини
D(+)-Биотин
0.013
D-калциев пантотенат
4,00
Холин хлорид
4,00
Фолиева киселина
4,00
мио-инозитол
7,20
Среди за клетъчни култури: Общ преглед
В областта на биологичните науки една от най-важните методики е клетъчната култура. Под „клетъчна култура“ се разбира извличането на клетки, тъкани или органи от животно или растение и последващото им имплантиране в изкуствена среда, благоприятна за тяхното оцеляване и/или растеж. Основните изисквания към средата за оптимално развитие на клетките са контролирана температура, субстрат за прикрепване на клетките, подходяща среда за растеж и инкубатор, който поддържа оптималното pH и осмолалност. Клетките трябва да разполагат с тези условия, за да могат да се развият до пълния си потенциал.
Изборът на подходяща среда за растеж за култивиране in vitro е етапът в клетъчната култура, който е едновременно най-критичен и най-важен. Средата за растеж, известна още като културална среда, е течност или гел, формулиран така, че да стимулира развитието на организмите на микроскопично, клетъчно или растително ниво. Средата, използвана за култивиране на клетки, често съдържа достатъчно количество енергия и вещества, които регулират клетъчния цикъл. Основните компоненти на културалната среда включват аминокиселини, витамини, неорганични соли, глюкоза и серум. Серумът се добавя към средата, тъй като действа като източник на растежни фактори, хормони и фактори за прикрепване. Освен че осигурява хранителни вещества, средата допринася и за поддържането на нивата на рН и осмолалността.
Видове среди, използвани в клетъчната култура
Както човешките, така и животинските клетки могат да се отглеждат както в изкуствена или синтетична среда, така и в изцяло естествена среда, допълнена с природни елементи. По-долу ще ви представим обзор на различните видове среди, които са налични към момента.
Естествени среди
В естествените среди се срещат само биологични течности, които съществуват в естественото си състояние. Естествените среди са много полезни и лесни за култивирането на широка гама от видове животински клетки. Липсата на разбиране за точните компоненти, съставляващи естествените среди, е основният фактор, допринасящ за ниската повторяемост на резултатите, получени при използването на естествени среди.
Изкуствени среди
Приготвянето на изкуствени или синтетични среди включва добавянето на хранителни вещества (както органични, така и неорганични), серумни протеини, въглехидрати, кофактори, витамини и соли, както и газови фази на O₂ и CO₂ [1].
Разработени са различни видове изкуствени среди, за да изпълняват една или повече от следните функции: 1) Незабавно оцеляване (балансиран солев разтвор с точно определено рН и осмотично налягане). 2) Продължително оцеляване (балансиран солев разтвор, допълнен с различни състави от органични химикали и/или серум). 3) Неограничено развитие. 4) Специализирани функции.
Съществуват четири различни класификации на изкуствените среди:
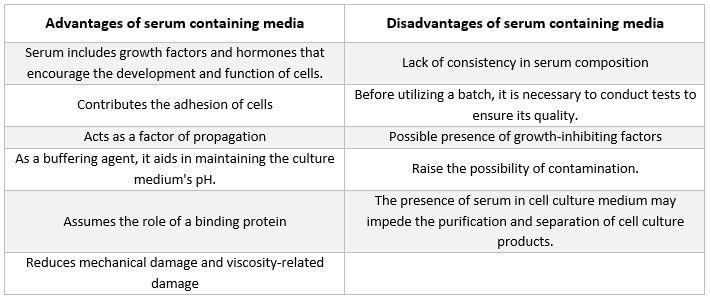
Среди, съдържащи серум
Най-често срещаният вид добавка в средите, използвани за отглеждане на животински клетки, е фетално телешко серум. То се добавя към културалната среда като евтина добавка, за да се постигнат възможно най-добрите условия за растеж. Освен че действа като преносител или хелатор на нестабилни или неразтворими във вода хранителни вещества, хормони и растежни фактори, инхибитори на протеази и други вещества, серумът също така свързва и неутрализира вредни молекули.
Среда без серум
Наличието на серум в средите има редица недостатъци и може да доведе до сериозни грешки при интерпретацията на резултатите в имунологичните изследвания [2, 3]. Създадени са различни видове безсерумни среди [4, 5]. Тези среди обикновено са специално формулирани, за да поддържат култивирането на един-единствен тип клетки, като например Knockout Serum Replacement и Knockout DMEM от Thermo Fisher Scientific, както и средата mTESR от Stem Cell Technologies [6], предназначена за стволови клетки [7].
Освен това тези среди съдържат определени количества пречистени растежни фактори, липопротеини и други протеини, които иначе обикновено се осигуряват от серума [8]. Тези среди често се наричат „дефинирани културални среди“, тъй като съставните им компоненти са добре изучени.
Химически дефинирани среди
Тези среди включват ултрачисти неорганични и органични компоненти, които не са били замърсени по никакъв начин. Те могат да включват и добавки от чисти протеини, като например растежни фактори.
Генетичната модификация на бактерии или дрожди, заедно с добавянето на определени мастни киселини, витамини, холестерол и аминокиселини, води до производството на техните съставни части [9].
Среди без протеини
Безпротеиновите среди са тези, които изобщо не съдържат протеини, а вместо това включват само непротеинови елементи. В сравнение със средите с добавен серум, използването на среди без добавени протеини спомага за по-голяма клетъчна пролиферация и протеинова експресия и улеснява пречистването на всеки продукт, получен в последващ процес [10–12]. Протеинът не се включва в формулировки като MEM и RPMI-1640. При необходимост обаче може да се добави протеинова добавка.
Културални среди и техните основни компоненти
Търговските културални среди могат да се закупят под формата на прах или течност и често съдържат разнообразни хранителни вещества, като аминокиселини, глюкоза, соли, витамини и други хранителни добавки.
Нуждите от тези компоненти са различни за всяка клетъчна линия, а тези различия са причината за голямото разнообразие от формулировки на среди. Всеки компонент отговаря за определена функция, която ще бъде описана в следващите параграфи:
Буферни системи
За да се поддържат оптимални условия за растеж, pH трябва да се контролира, което често се извършва чрез една от двете буферни системи:
Естествена буферна система
Съотношението CO₂/H₂CO₃ в атмосферата е равно на това в средата, което създава естествен буферен механизъм. За да се запази този естествен буферен механизъм, културите трябва да се поддържат във въздушна среда с 5–10% CO₂, което често се постига чрез използване на CO₂ инкубатор. Едно от най-големите предимства на използването на естествен буфер е, че той е евтин и безопасен.
HEPES
Химичното буфериране с помощта на цвитериона HEPES има по-голяма буферна способност в диапазона на pH 7,2–7,4 и не изисква регулирана газова среда. За определени видове клетки по-голяма доза HEPES може да бъде вредна. Средите, съдържащи HEPES, са също така много по-податливи на фототоксичните ефекти на флуоресцентната светлина [13].
Фенол червено
Индикаторът за рН фенол червено често се включва в търговски достъпните културални среди, което позволява непрекъснато наблюдение на рН. При размножаването на клетките метаболитите, произвеждани от тях, предизвикват промяна в рН и съответно промяна в цвета на средата. Фенол червено оказва двоен ефект върху цвета на средата, като я оцветява в жълто при кисело pH и в лилаво при алкално pH. pH 7,4, оптималната стойност за клетъчна култура, кара средата да изглежда флуоресцентно червена.
Фенол червено обаче има няколко недостатъка: Първо, фенол червено може да имитира действието на редица стероидни хормони, предимно естроген [14]. Поради това при изследване на естроген-чувствителни клетки, като например тъкан от млечната жлеза, се препоръчва среда без фенол червено. Натриево-калиевият баланс се нарушава от присъствието на фенол червено в няколко безсерумни формулировки. Добавянето на серум или говежди хипофизен хормон към средите може да неутрализира този ефект [15]. На трето място, откриването при експерименти с проточна цитометрия се затруднява от присъствието на фенол червено.
Неорганични соли
Средите, съдържащи неорганични соли, като натриеви, калиеви и калциеви йони, спомагат за поддържането на осмотичното равновесие и регулират мембранния потенциал.
Аминокиселини
Тъй като аминокиселините са основните съставни части на протеините, те са незаменим компонент на всяка културална среда за клетъчен растеж, която някога е била създадена. Тъй като клетките не са в състояние сами да произвеждат определени аминокиселини, е важно културалната среда да съдържа незаменими аминокиселини. Те са необходими за пролиферацията на клетките, а концентрацията, в която се съдържат, определя максималната клетъчна плътност, която може да бъде постигната. По-специално, L-глутаминът, една незаменима аминокиселина, е от особено голямо значение.
L-глутаминът функционира като вторичен източник на енергия за метаболизма и доставя азот за производството на NAD, NADPH и нуклеотиди. Тъй като L-глутаминът е нестабилна аминокиселина, която с времето се превръща във форма, която клетките не могат да усвоят, той трябва да се добавя към средата.
Освен това в средата могат да се добавят неесенциални аминокиселини, за да се възстановят тези, които са били изразходвани по време на процеса на растеж. Растежът на клетките се ускорява, а жизнеспособността им се повишава, когато средата за растеж се допълва с неесенциални аминокиселини.
Въглехидрати
Въглехидратите под формата на захари са основният източник на енергия. Много от средите съдържат също малтоза и фруктоза, в допълнение към по-разпространените захари – глюкоза и галактоза.
Протеини и пептиди
Албуминът, трансферинът и фибронектинът са най-често използваните протеини и пептиди. Те са особено важни в среди, които не съдържат серум. Албуминът, трансферинът, апротининът, фетуинът и фибронектинът са някои от протеините, които могат да се открият в серума, който е богат източник на протеини.
Албуминът е основният протеин в кръвта и неговата функция е да свързва и транспортира различни вещества, включително вода, соли, свободни мастни киселини, хормони и витамини, между различните органи и клетки. Способността на албумина да се свързва с химични вещества го прави ефективен кандидат за отстраняване на вредни съединения от средата, в която се култивират клетките.
Апротининът е защитен агент в системите за клетъчна култура, тъй като е стабилен при неутрално и кисело рН, както и устойчив на високи температури и на разрушаването, което може да бъде причинено от протеолитични ензими. Той е способен да инхибира редица серинови протеази, включително трипсин, наред с други.
Фетуинът е гликопротеин, който може да бъде открит в по-големи количества в серума на плода и новородените животни в сравнение със серума на възрастните. В допълнение към това той действа като инхибитор на сериновите протеази. Протеинът фибронектин е основен компонент в процеса на клетъчната адхезия. Трансферинът е протеин, който транспортира желязо и отговаря за доставянето на желязо до клетъчните мембрани.
Мастни киселини и липиди
Те играят решаваща роля в безсерумната среда, когато липсва серум.
Витамини
Многобройни витамини са необходими за развитието и пролиферацията на клетките. Витамините не могат да се произвеждат в достатъчни количества от клетките и затова са незаменими в тъканните култури като хранителни добавки.
При клетъчната култура серумът е основният източник на витамини; обаче средите се обогатяват и с различни витамини, за да бъдат подходящи за конкретен тип клетки. Най-често витамините от група В се използват за стимулиране на растежа.
Микроелементи
Химични елементи като мед, цинк, селен и междинни съединения на трикарбоксилната киселина са известни като микроелементи. Микроелементите често се добавят към среди, които не съдържат серум, за да заменят тези, които обикновено се съдържат в серума. Тези елементи са важни химични компоненти, необходими за здравословното развитие на клетките. Много биохимични реакции зависят от определени микроелементи, като например ензимната активност.
Добавки към средата
Пълната среда за растеж, препоръчвана за определени клетъчни линии, се нуждае от допълнителни компоненти, които липсват в базовите среди и серума. Тези хранителни добавки подпомагат клетъчния растеж и нормалната метаболитна функция.
Въпреки че хормоните, растежните фактори и сигналните молекули са от съществено значение за правилното размножаване на конкретни клетъчни линии, винаги трябва да се спазват следните предпазни мерки: Тъй като добавянето на добавки може да промени осмолалността на пълната среда за растеж, което може да попречи на развитието на клетките, винаги е препоръчително да се провери осмолалността след добавянето на добавки. За по-голямата част от клетъчните линии оптималната осмолалност варира между 260 и 320 mOSM/kg.
Антибиотици
Антибиотиците често се използват за потискане на развитието на бактериални и гъбични замърсители [16], въпреки че не са необходими за клетъчния растеж. Тъй като антибиотиците могат да прикрият замърсяване с микоплазма и резистентни бактерии, рутинното им използване не се препоръчва при клетъчни култури [17, 18].
Освен това антибиотиците могат да нарушат метаболизма на свръхчувствителните клетки. Често се използват комбинациите от пеницилин и стрептомицин, произведени от MilliporeSigma и Life Technologies. Плазмоцин е бил използван при култивирането на глиомните клетъчни линии TS603, TS516 и BT260 [19] и е доказано, че е ефективен при отстраняването на микоплазмена контаминация (20).
Серум
В серума се съдържат албумини, растежни фактори и инхибитори на растежа. Серумът е един от най-важните компоненти на средата за клетъчна култура, тъй като осигурява аминокиселини, протеини, витамини (особено мастноразтворими витамини като А, D, Е и К), въглехидрати, липиди, хормони, растежни фактори, минерали и микроелементи.
Често се използва серум от фетални и телешки източници, за да се стимулира развитието на култивираните клетки. Феталният серум е богат източник на растежни фактори и е подходящ за клониране на клетки и развитието на чувствителни клетки. Поради намалените си способности за стимулиране на растежа телешкият серум се използва в експерименти за контактна инхибиция. Обикновено културалните среди съдържат от 2% до 10% серум. Добавянето на серум към културалната среда служи за следните цели [21]:
-
Серумът доставя необходимите за клетките хранителни вещества (както в разтвор, така и свързани с протеини).
-
В серума са включени няколко растежни фактори и хормони, участващи в стимулирането на растежа и специализираната клетъчна активност.
-
То предоставя много свързващи протеини, като албумин и трансферин, които транспортират други химични вещества в клетката. Например, албуминът доставя мазнини, витамини, хормони и др. в клетките.
-
То осигурява също така протеини, като фибронектин, които увеличават клетъчната адхезия към субстрата. Освен това, то произвежда елементи за разпространение, които подпомагат разширяването на клетките преди деленето.
-
То доставя инхибитори на протеазите, които предотвратяват протеолизата в клетките.
-
Съдържа също минерали като Na+, K+, Zn2+ и Fe2+.
-
Той повишава вискозитета на средата, като по този начин предпазва клетките от механични увреждания по време на разбъркването на суспензионната култура.
-
Той действа и като буфер.
Източници
[1] Morgan J, Morton H, Parker R. Хранене на животински клетки в тъканна култура; първоначални проучвания върху синтетична среда. Proc Soc Exp Biol Med. 1950;73:1-8
[2] Kerbel R, Blakeslee D. Бърза адсорбция на компонент от фетално телешко серум от клетки на бозайници в култура. Потенциален източник на артефакти в проучванията на антисера срещу клетъчно-специфични антигени. Immunology. 1976;31:881-91
[3] Сула К., Драбер П., Нуза К. Добавянето на серум към средата, използвана за приготвяне на клетъчни суспензии, като възможен източник на артефакти при клетъчно-медиирани реакции, изследвани посредством теста с поплитеалния лимфен възел. J Immunogenet. 1980;7:483-9
[4] Мариани Е., Мариани А., Монако М., Лали Е., Витале М., Факини А. Търговски безсерумни среди: растеж на хибридоми и производство на моноклонални антитела. J Immunol Methods. 1991;145:175-83
[5] Barnes D, Sato G. Методи за отглеждане на култивирани клетки в безсерумна среда. Anal Biochem. 1980;102:255-70
[6] Ю Х., Лу С., Гасиор К., Сингх Д., Васкес Санчес С., Тапиа О. и др. Шапероните HSP70 пренасят TDP-43 без РНК в анизотропни интрануклеарни течни сферични обвивки. Science. 2021;371:
[7] Meharena H, Marco A, Dileep V, Lockshin E, Akatsu G, Mullahoo J, и др. Стареенето, предизвикано от синдрома на Даун, нарушава ядрената архитектура на невронните предшественици. Cell Stem Cell. 2022;29:116-130.e7
[8] Iscove N, Melchers F. Пълно заместване на серума с албумин, трансферин и соеви липиди в култури от В-лимфоцити, реагиращи на липополизахариди. J Exp Med. 1978;147:923-33
[9] Stoll T, Muhlethaler K, von Stockar U, Marison I. Систематично усъвършенстване на химически дефинирана среда без протеини за растеж на хибридоми и производство на моноклонални антитела. J Biotechnol. 1996;45:111-23
[10] Darfler F. Среда без протеини за растеж на хибридоми и други клетки на имунната система. In Vitro Cell Dev Biol. 1990;26:769-78
[11] Barnes D, Sato G. Клетъчна култура без серум: обединяващ подход. Cell. 1980;22:649-55
[12] Hamilton W, Ham R. Клонален растеж на клетъчни линии от китайски хамстер в безпротеинови среди. In Vitro. 1977;13:537-47
[13] Zigler J, Lepe Zuniga J, Vistica B, Gery I. Анализ на цитотоксичните ефекти на изложена на светлина културална среда, съдържаща HEPES. In Vitro Cell Dev Biol. 1985;21:282-7
[14] Бертоа И., Каценеленбоген Ж., Каценеленбоген Б. Фенол червено в средите за тъканни култури е слаб естроген: последствия за изследването на естроген-чувствителни клетки в култура. Proc Natl Acad Sci U S A. 1986;83:2496-500
[15] Karmiol S. Разработване на безсерумни среди. В: Master JRW, редактор. Клетъчна култура на животни, 3-то изд. Оксфорд: Oxford University Press; 2000.
[16] Perlman D. Използване на антибиотици в среди за клетъчни култури. Methods Enzymol. 1979;58:110-6
[17] McGarrity G. Разпространение и контрол на микоплазмена инфекция в клетъчни култури. In Vitro. 1976;12:643-8
[18] Мастърс Дж., Стейси Г. Смяна на средата и пасажиране на клетъчни линии. Nat Protoc. 2007;2:2276-84
[19] Chakraborty A, Laukka T, Myllykoski M, Ringel A, Booker M, Tolstorukov M и др. Хистоновата деметилаза KDM6A директно усеща кислорода, за да контролира хроматина и съдбата на клетката. Science. 2019;363:1217-1222
[20] Мола Каземиха В., Азари С., Аманзаде А., Бонакдар С., Шоджаеи Могадам М., Хабиби Анбухи М. и др. Ефективност на Plasmocin™ върху различни клетъчни линии на бозайници, инфектирани с моликути, в сравнение с често използвани антибиотици в клетъчни култури: местен опит. Cytotechnology. 2011;63:609-20
[21] Краг Хансен У. Молекулни аспекти на свързването на лиганда със серумния албумин. Pharmacol Rev. 1981;33:17-53