
خلايا HepG2 - مورد لأبحاث سرطان الكبد
Hep-G2 هي سلالة خلايا سرطان الكبد البشري التي نشأت من نسيج الكبد لرجل قوقازي يبلغ من العمر 15 عامًا مصاب بسرطان الخلايا الكبدية. تُستخدم هذه الخلايا بشكل متكرر في دراسات استقلاب الأدوية والسمية الكبدية. على الرغم من أن خلايا HepG2 تتميز بمعدلات تكاثر عالية ومظهر شبيه بالخلايا الظهارية، إلا أنها غير مسببة للأورام وتؤدي وظائف كبدية متنوعة ومتمايزة. في عام 1975، استخلص الباحثون خلايا HepG2 من سرطان الخلايا الكبدية، مما جعلها أول سلالة خلوية كبدية تظهر الخصائص الحاسمة للخلايا الكبدية. على عكس خط الخلايا SK-Hep1 الذي تم إنشاؤه سابقًا، والذي يفتقر إلى علامات خلايا الكبد الأساسية، يمكن لخلايا HepG2 إفراز بروتينات بلازما متنوعة وتوفر نموذجًا قيمًا لدراسة الديناميكيات داخل الخلايا لمجالات سطح الخلية في خلايا الكبد البشرية. تظهر هذه الخلايا شكلاً شبيهاً بالخلايا الظهارية، ويبلغ عدد كروموسوماتها النموذجي 55، ويمكن تحفيزها بهرمون النمو البشري.
- وسط النمو
- انظر صفحة المنتج
- زمن التضاعف
- انظر صفحة المنتج
- نوع النمو
- ملتصق
- مستوى السلامة البيولوجية
- BSL-1
- متوفر من
- Cytion — طلب HepG2
خصائص خلايا HepG2
الشكل النموذجي لخلايا الكبد الأولية هو مكعب وتحتوي عادة على نواتين. في المقابل، تتميز خلايا HepG2 بمورفولوجيا شبيهة بالظهارة مع نواة واحدة وعدد كروموسومات يتراوح بين 48 و54 لكل خلية. على الرغم من أن خلايا HepG2 يمكن أن تمثل ما يصل إلى 25٪ من إجمالي البروتين الخلوي، إلا أن حجمها أكبر من حجم خلايا الكبد الطبيعية، حيث تشكل حوالي 10٪ من البروتين الكامل في الخلية. تلعب البروتينات الخلوية دورًا حاسمًا داخل الخلية، حيث تؤدي الوظائف المحددة بواسطة الجينات.
غالبًا ما تظهر الخلايا السرطانية، بما في ذلك تلك التي تحتوي على عدد غير طبيعي من الكروموسومات، زيادة في عدد النوى، تصل إلى سبعة نوى لكل خلية. نظرًا لدرجة التمايز العالية في المختبر، توفر خلايا HepG2 نموذجًا مثاليًا لدراسة النقل داخل الخلايا وديناميكيات القنوات الصفراوية، وبروتينات الغشاء الجيبي، والدهون في خلايا الكبد البشرية.
يبلغ متوسط قطر خلية HepG2 حوالي 10-20 ميكرومتر، وهو أصغر من خلايا الكبد التي يبلغ قطرها 15 ميكرومتر، ولكنه مشابه لخلايا الأورام المصابة بورم الكبد (HB)، والتي يتراوح قطرها بين 10-20 ميكرومتر.
علم الوراثة لخلايا HepG2
يُظهر خط الخلايا Hep-G2 عدة انتقالات، بما في ذلك تلك التي تحدث بين الأذرع القصيرة للكروموسومات 1 و21، وتثلث الصبغيات للكروموسومات 2 و16 و17، ورباعية الصبغيات للكروموسوم 20. كما لوحظ فقدان منطقة الكروموسوم 4q3، المرتبطة بالانتقال t(1;4) الذي غالبًا ما يُرى في ورم الكبد (HB) وتشوهات كروموسومية أخرى، مثل ثلاثيات الكروموسومات 2 و20. يتراوح عدد الكروموسومات في خلايا HepG2 من 50 إلى 60، مما يشير إلى نمط كروموسومي مفرط ثنائي الصيغة الصبغية، في حين تظهر بعض الحالات أكثر من 100 كروموسوم وتتميز بتضخم رباعي الصيغة الصبغية. تحتوي خلايا HepG2 على حوالي 7.5 بيكوغرام من الحمض النووي، أي 15٪ أكثر من الخلية الجسدية المتوسطة. وبالمقارنة، فإن خلايا الكبد الأولية لها شكل خلية مكعبة وتحتوي عادةً على نواتين [1].
الملف الجيني لخلايا HepG2
يحمل خط الخلايا HepG2 طفرة C228T في منطقة محفز TERT، وهي موجودة أيضًا في سرطان الخلايا الكبدية (HCC) وورم الكبد (HB). تساهم هذه الطفرة في الخلود عن طريق حماية التيلوميرات في الخلايا السرطانية. بالإضافة إلى ذلك، تُظهر خلايا HepG2 جين TP53 من النوع البري، وهو جين حاسم في قمع السرطان البشري، حيث يلعب دورًا في توقف دورة الخلية، وموت الخلايا المبرمج، والشيخوخة. يمكن أن تؤدي الطفرات في هذا الجين إلى تعزيز تكاثر الخلايا.
تشارك خلايا HepG2 في عدة مسارات، بما في ذلك خلل تنظيم نمو الخلايا، ومسارات البقاء مثل ورم الكبد الجنيني والجنيني، ومسار Wnt/β-catenin. علاوة على ذلك، تتميز هذه السلالة الخلوية بحذف مميز للإكسون الثالث من جين CTNNB1، وهو مطابق لما يُلاحظ في سرطان الكبد من النوع الظهاري [2،3].

نظرة عامة على خلايا سرطان الخلايا الكبدية HepG2 في أبحاث الكبد
أصبحت خلايا HepG2، التي تنشأ من ورم الكبد البشري، أداة لا تقدر بثمن في أبحاث وظائف الكبد وأمراضه، بما في ذلك سرطان الخلايا الكبدية. توفر هذه السلالات الخلوية الكبدية رؤى ثاقبة حول الاستجابات الخلوية لخلايا الكبد البشرية في ظل ظروف تجريبية متنوعة. وقد كان استخدام البلازميدات المبلغة عن اللوسيفيراز في خلايا HepG2 فعالاً بشكل خاص في تتبع التعبير الجيني والتحويلات الخلوية، والتي تعتبر أساسية في أبحاث التمثيل الغذائي، مثل دراسة تأثيرات الإيثانول على خلايا الكبد.
دراسات العدوى الفيروسية وأمراض الكبد باستخدام خلايا HepG2
تعد سلالات الخلايا السرطانية الكبدية المُخلدة مثل HepG2 و Huh7 ضرورية في دراسة العدوى الفيروسية، حيث تُظهر تكرار دورة الخلية الكاملة لفيروس التهاب الكبد D (HDV) والتعبير الجيني لفيروس التهاب الكبد B (HBV) [5،6]. وبالتوازي مع ذلك، تلعب سلالات خلايا HepaRG دورًا حاسمًا في توضيح آليات دخول فيروس التهاب الكبد B [7]. كما تُستخدم خلايا HepG2 لدراسة مجموعة متنوعة من أمراض الكبد البشرية، بدءًا من الحالات الوراثية مثل الركود الصفراوي داخل الكبد العائلي التقدمي (PFIC) ومتلازمة دوبين-جونسون وصولًا إلى الدراسات البيئية والغذائية المتعلقة بالعوامل السامة للخلايا والعوامل السامة للجينات، وكذلك في استهداف الأدوية وأبحاث تكوين سرطان الكبد [8,9]. يمتد استخدامها إلى التجارب التي تُجرى باستخدام أجهزة الكبد الاصطناعية الحيوية.
تفاعلات خلايا HepG2 مع المواد الحيوية في هندسة الأنسجة
يعد تفاعل خلايا HepG2 مع المواد الحيوية المختلفة أمرًا محوريًا في هندسة الأنسجة. تساعد تقنيات مثل تقنية المسبار الغرواني على فهم هذه التفاعلات من خلال قياس خصائص الالتصاق الخلوي، والتي تعتبر حيوية في تحديد قابلية الخلايا للحياة من أجل تطوير هياكل داعمة ونماذج دقيقة لأنسجة الكبد.
سلوك الخلايا والابتكارات في النماذج القائمة على HepG2
تعد دراسة سلوك الخلايا في النماذج القائمة على HepG2 أمرًا بالغ الأهمية لأبحاث أمراض الكبد. وقد أدى التقدم في مزارع الخلايا الكروية ثلاثية الأبعاد إلى إنشاء كريات خلايا HepG2، مما يوفر نموذجًا أكثر صلة من الناحية الفسيولوجية يعكس بشكل وثيق خلايا الكبد الطبيعية. تشير هذه النماذج ثلاثية الأبعاد، مع زيادة النشاط الأيضي، إلى إمكانية استخدام خلايا HepG2 كنموذج لورم الكبد، وهي مهمة في أبحاث علاج السرطان، خاصة لمحاكاة أورام الكبد واختبار الأساليب العلاجية الجديدة [10-12].
مقارنة وخصائص HepG2 بين سلالات الخلايا السرطانية الأخرى
تعد HepG2 واحدة من أكثر سلالات الخلايا السرطانية الكبدية استخدامًا، وقد تم اختيارها لتطبيقاتها الواسعة في البحث العلمي من بين حوالي 40 سلالة خلية سرطانية كبدية متاحة [13]. على الرغم من ضعف أو غياب التعبير عن بعض إنزيمات السيتوكروم P450 مقارنة بالخلايا الكبدية الطبيعية، فقد دفع الملف الأيضي لـ HepG2 الجهود لتعديل خط الخلايا من أجل دراسات أفضل لاستقلاب الأدوية [13]. بالمقارنة مع سلالات الخلايا السرطانية مثل MCF7 و PC3 و 143B و HEK293، تُظهر خلايا HepG2 ملامح فريدة لمحتوى الأحماض الأمينية التي تؤثر بشكل كبير على تخليق البروتين وإفرازه، مما يسلط الضوء على مساراتها الأيضية الفريدة [14].
استكشاف أبحاث أمراض الكبد باستخدام خلايا HepG2
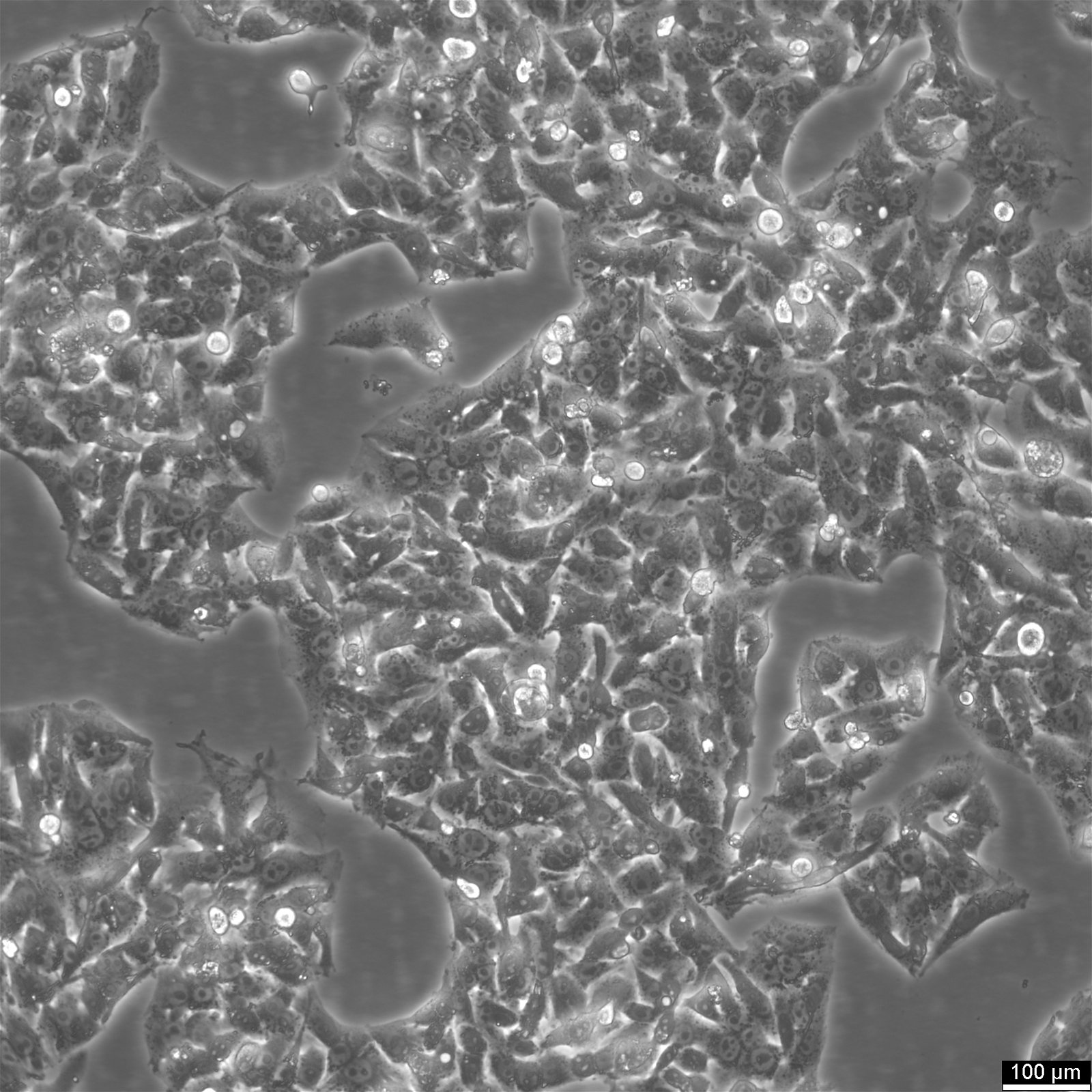
إعادة زراعة خلايا HepG2
فيما يلي خمس خطوات لإزالة الخلايا الملتصقة من قوارير زراعة الخلايا باستخدام Accutase:
- أزل الوسط من قارورة زراعة الخلايا واشطف الخلايا الملتصقة باستخدام PBS بدون الكالسيوم والمغنيسيوم. استخدم 3-5 مل من PBS لقوارير T25 و5-10 مل لقوارير T75.
- أضف Accutase إلى قارورة زراعة الخلايا، باستخدام 1-2 مل لكل قارورة T25 و2.5 مل لكل قارورة T75. تأكد من أن Accutase يغطي طبقة الخلايا بالكامل.
- قم بتحضين القارورة في درجة حرارة الغرفة لمدة 8-10 دقائق.
- أعد تعليق الخلايا بعناية باستخدام الوسط، باستخدام 10 مل من الوسط الطازج.
- قم بفصل الخلايا المعاد تعليقها في جهاز الطرد المركزي لمدة 5 دقائق عند 300xg، ثم أعد تعليقها في وسط جديد، وقم بتوزيعها في قوارير جديدة تحتوي على وسط جديد.
الآفاق المستقبلية لخلايا HepG2
يستمر السعي لإطلاق العنان للإمكانات الكاملة لسلسلة خلايا HepG2 مع التقدم الرائد في زيادة تعبير السيتوكروم. كما يستكشف الباحثون إمكانية زراعة الخلايا الكروية ثلاثية الأبعاد، والتي توفر نظامًا أكثر صلة من الناحية الفسيولوجية. النشاط الأيضي، بما في ذلك السيتوكرومات، أعلى بشكل ملحوظ في نماذج HepG2 الكروية ثلاثية الأبعاد مقارنة بالخلايا ثنائية الأبعاد، مما يقربنا من إنشاء نموذج يعكس خلايا الكبد الطبيعية. بالإضافة إلى ذلك، يمكن أن يمهد استكشاف العمليات الديناميكية الكامنة وراء التوزيع غير الصحيح لبروتينات سطح الخلية الطريق لفهم أفضل لأمراض الكبد.
خلايا HepG2: فهم دورها وخصائصها المميزة في مجال البحوث الطبية الحيوية - الأسئلة الشائعة
المراجع
- Vyas, R.C., Darroudi, F., Natarajan, A.T. Radiation-induced chromosomal breakage and rejoining in interphase-metaphase chromosomes of human lymphocytes, Mutat Res, 1991; 249(1):29-35.
- Woodfield, S.E., Shi, Y., Patel, R.H., Chen, Z., Shah, A.P., Srivastava, R.K., Whitlock, R.S., Ibarra, A.M., Larson, S.R., سارابيا، س. ف.، وآخرون. تثبيط MDM4: استراتيجية علاجية جديدة لإعادة تنشيط P53 في ورم الكبد. Sci. Rep. 2021، 11، 2967.
- Hussain, S.P., Schwank, J., Staib, F., Wang, X.W., Harris, C.C. طفرات TP53 وسرطان الخلايا الكبدية: رؤى حول مسببات وتطور سرطان الكبد. Oncogene 2004.
- شيت، ج.، سايدمان، ل.، هانسل، ر.، سيهوفر، د.، دام، ج. دراسة نقدية لقابلية استخدام خطوط الخلايا السرطانية الكبدية HepG2 و Huh7 كنماذج للتمثيل الأيضي لسرطان الخلايا الكبدية القابل للاستئصال. Cancers 2022, 14(17), 4227.
- Verrier, E.R., Colpitts, C.C., Schuster, C., Zeisel, M.B., Baumert, T.F. نماذج زراعة الخلايا لدراسة عدوى فيروسات التهاب الكبد B و D. Viruses 2016, 8, 261.
- Verrier, E.R., Colpitts, C.C., Bach, C., Heydmann, L., Weiss, A., Renaud, M., Durand, S.C., Habersetzer, F., Durantel, D., أبو جاودي، ج.، وآخرون. فحص موجه للتداخل الوظيفي للـ RNA يكشف عن الجليبيكان 5 كعامل دخول لفيروسات التهاب الكبد B و D. Hepatology 2016, 63, 35–48.
- Gripon, P., Rumin, S., Urban, S., Le Seyec, J., Glaise, D., Cannie, I., Guyomard, C., Lucas, J., Trepo, C., Guguen-Guillouzo, C. إصابة خط خلايا سرطان الكبد البشري بفيروس التهاب الكبد B. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660.
- Mersch-Sundermann, V., Knasmüller, S., Wu, X.J., Darroudi, F., Kassie, F. استخدام خط خلايا كبدية مشتقة من الإنسان للكشف عن العوامل الحامية للخلايا والمضادة للتسمم الجيني والمضادة للتسمم الجيني المشترك. علم السموم. 2004؛ 198(1–3): 329–340.
- فانيلي، أ. HepG2 (سرطان الخلايا الكبدية): زراعة الخلايا. HepG2. تم الاسترجاع في 3 ديسمبر 2017.
- Xuan, J., Chen, S., Ning, B., Tolleson, W.H., Guo, L. تطوير خلايا مشتقة من HepG2 تعبر عن السيتوكروم P450s لتقييم سمية الكبد المرتبطة بالأيض والمستحثة بالأدوية. Physiol. Behav. 2017, 176, 139–148.
- Ooka, M., Lynch, C., Xia, M. تطبيق تنشيط الأيض في المختبر في الفحص عالي الإنتاجية. Int. J. Mol. Sci. 2020, 21, 8182.
- هوانغ، ل.، كوفتري، م. و. هـ.، هسو، هـ. انخفاض تنظيم جين ديهيدروإيباندروستيرون سلفوترانزفيراز في سرطان الخلايا الكبدية البشري. Mol. Cell. Endocrinol.
- Zhu, Z., Hao, X., Yan, M., et al. الخلايا الجذعية/السلفية السرطانية غنية جدًا في مجموعة CD133 + CD44 + في سرطان الخلايا الكبدية. Int J Cancer. 2010; 126:2067-2078.
- Arbus, C., Benyamina, A., Llorca, P.-M., Baylé, F., Bromet, N., Massiere, F., Garay, R.P., Hameg, A. توصيف إنزيمات السيتوكروم P450 البشرية المشاركة في استقلاب السياميمازين. Eur J Pharm Sci. ديسمبر 2007;32(4-5):357-66.