
HepG2 sejtek – a májrák kutatásának forrása
A Hep-G2 egy humán májrákos sejtvonal, amely egy 15 éves, hepatocelluláris karcinómában szenvedő kaukázusi férfi májszövetéből származik. Ezeket a sejteket gyakran használják gyógyszer-metabolizmus és hepatotoxicitás vizsgálatokban. Bár a HepG2 sejtek szaporodási aránya magas és megjelenésük hámsejtszerű, nem tumorigenikusak és különböző differenciált májfunkciókat látnak el. 1975-ben a kutatók hepatocelluláris karcinómából nyerték ki a HepG2 sejteket, így ez lett az első májsejtvonal, amely a hepatociták kritikus jellemzőit mutatta. A korábban létrehozott SK-Hep1 sejtvonal ellentétben, amelyből hiányoznak az alapvető májsejt-markerek, a HepG2 sejtek különböző plazmafehérjéket képesek kiválasztani, és értékes modellt nyújtanak az emberi hepatociták sejtfelszíni doménjeinek intracelluláris dinamikájának tanulmányozásához. Ezek a sejtek epitheliális jellegű morfológiát mutatnak, kromoszómaszámuk általában 55, és emberi növekedési hormonnal stimulálhatók.
- Növekedési tápközeg
- Lásd a termékoldalt
- Duplázódási idő
- Lásd a termékoldalt
- Növekedési típus
- Adherens
- Biológiai biztonsági szint
- BSL-1
- Kapható
- Cytion — HepG2 megrendelése
A HepG2 sejtek jellemzői
Az elsődleges hepatociták jellegzetes alakja kocka alakú, és általában két sejtmagot tartalmaznak. Ezzel szemben a HepG2 sejtek epitheliális jellegű morfológiával rendelkeznek, egyetlen sejtmaggal és sejtenként 48–54 kromoszómával. Bár a HepG2 sejtek a teljes sejtfehérje akár 25%-át is kitehetik, méretük nagyobb, mint a normál hepatocitáké, és a sejt teljes fehérjéjének körülbelül 10%-át teszik ki. A sejtfehérjék kritikus szerepet játszanak a sejten belül, végrehajtva a gének által meghatározott funkciókat.
A tumorsejtek, beleértve a rendellenes kromoszómaszámúakat is, gyakran mutatnak megnövekedett sejtmagszámot, akár hét sejtmagot is elérve sejtenként. In vitro magas differenciáltságuknak köszönhetően a HepG2 sejtek ideális modellt nyújtanak az emberi hepatocitákban az epevezetékek, a szinuszos membránfehérjék és a lipidek intracelluláris transzportjának és dinamikájának tanulmányozásához.
A HepG2 sejtek átlagos átmérője körülbelül 10–20 µm, ami kisebb, mint a 15 µm átmérőjű hepatocitáké, de hasonló a 10–20 µm közötti átmérőjű hepatoblastoma (HB) tumorsejtekéhez.
A HepG2 genetikája
A Hep-G2 sejtvonal számos transzlokációt mutat, beleértve a 1. és 21. kromoszóma rövid karjai közötti transzlokációkat, a 2., 16. és 17. kromoszóma triszómiáit, valamint a 20. kromoszóma tetraszómiáját. Megfigyelhető a 4q3 kromoszómarégió elvesztése is, amely a hepatoblastomában (HB) gyakran előforduló t(1;4) transzlokációval és más kromoszómaabnormitásokkal, például a 2. és 20. kromoszóma triszómiájával társul. A HepG2 sejtek kromoszómaszáma 50 és 60 között mozog, ami hiperdiploid kariotípust jelez, míg egyes esetekben több mint 100 kromoszóma figyelhető meg, és ezeket tetraploid megnagyobbodás jellemzi. A HepG2 sejtek körülbelül 7,5 pg DNS-t tartalmaznak, ami 15%-kal több, mint egy átlagos szomatikus sejtben. Ehhez képest az elsődleges hepatociták sejtjei kocka alakúak és jellemzően két sejtmagot tartalmaznak [1].
A HepG2 sejtek mutációs profilja
A HepG2 sejtvonal hordozza a TERT promóter régió C228T mutációját, amely a hepatocelluláris karcinómában (HCC) és a hepatoblastómában (HB) is jelen van. Ez a mutáció hozzájárul az immortalisációhoz azáltal, hogy védi a ráksejtek telomereit. Ezenkívül a HepG2 sejtek vad típusú TP53-at mutatnak, amely kritikus gén az emberi rák szuppressziójában, mivel szerepet játszik a sejtciklus leállásában, az apoptózisban és az öregedésben. E gén mutációi elősegíthetik a sejtek proliferációját.
A HepG2 sejtek számos útvonalban részt vesznek, beleértve a sejtnövekedés szabályozási zavarait, a túlélési útvonalakat, mint például a magzati és embrionális HB, valamint a Wnt/β-katenin útvonalat. Ezenkívül a sejtvonalban jellegzetes a CTNNB1 gén harmadik exonjának deléciója, amely megegyezik az epiteliális típusú HB-ben megfigyeltel [2,3].

A HepG2 májsejtes karcinóma sejtek áttekintése a májkutatásban
Az emberi hepatómából származó HepG2 sejtek felbecsülhetetlen értékű eszközzé váltak a májfunkciók és -betegségek, köztük a hepatocelluláris karcinóma kutatásában. Ezek a májsejtvonalak betekintést nyújtanak az emberi májsejtek sejtválaszaiba különböző kísérleti körülmények között. A luciferáz riporter plazmidok használata HepG2 sejtekben különösen hatékonynak bizonyult a génexpresszió és a sejttranszfekciók nyomon követésében, amelyek alapvető fontosságúak az anyagcsere-kutatásban, például az etanol májsejtekre gyakorolt hatásainak vizsgálatában.
Vírusfertőzések és májbetegségek vizsgálata HepG2 sejtek felhasználásával
Az olyan halhatatlanná tett májdaganat-sejtvonalak, mint a HepG2 és a Huh7, elengedhetetlenek a vírusfertőzések tanulmányozásában, mivel bizonyítják a hepatitis D (HDV) teljes sejtciklus-replikációját és a hepatitis B (HBV) expresszióját [5,6]. Ezzel párhuzamosan a HepaRG sejtvonalak kritikus szerepet játszanak a HBV behatolási mechanizmusainak tisztázásában [7]. A HepG2 sejteket számos emberi májbetegség vizsgálatára is alkalmazzák, a progresszív familiáris intrahepatikus cholestasis (PFIC) és a Dubin-Johnson-szindróma genetikai állapotaitól kezdve a citotoxikus és genotoxikus anyagokkal kapcsolatos környezeti és táplálkozási vizsgálatokig, valamint a gyógyszercélzás és a hepatokarcinogenezis kutatásában [8,9]. Használatuk kiterjed a bio-mesterséges máj eszközökkel végzett kísérletekre is.
A HepG2 sejtek és a biológiai anyagok kölcsönhatása a szövetmérnökségben
A HepG2 sejtek és a különböző biológiai anyagok közötti kölcsönhatás kulcsfontosságú a szövetmérnökségben. Az olyan technikák, mint a kolloid szondás technika, segítenek megérteni ezeket a kölcsönhatásokat a sejtadhéziós tulajdonságok mérésével, amelyek elengedhetetlenek a sejtek életképességének meghatározásához a vázszerkezetek és a pontos májszövet-modellek fejlesztése érdekében.
Sejtviselkedés és innovációk a HepG2-alapú modellekben
A HepG2-alapú modellekben a sejtek viselkedésének tanulmányozása elengedhetetlen a májbetegségek kutatásához. A háromdimenziós szferoid sejtkultúrák terén elért fejlődés a HepG2 sejt szferoidok létrehozásához vezetett, amelyek egy fiziológiailag relevánsabb modellt kínálnak, amely szorosan tükrözi a normális hepatocitákat. Ezek a megnövekedett metabolikus aktivitású 3D-modellek arra utalnak, hogy a HepG2-sejtek alkalmasak lehetnek hepatoblastoma modellként, és jelentős szerepet játszanak a rákkezelés kutatásában, különösen a májdaganatok szimulálásában és új terápiás megközelítések tesztelésében [10-12].
A HepG2 összehasonlítása és jellemzői más tumorsejtvonalakkal
A HepG2 az egyik legszélesebb körben használt májdaganatos sejtvonal, amelyet a mintegy 40 rendelkezésre álló májdaganatos sejtvonal közül választottak ki széles körű tudományos kutatási alkalmazásai miatt [13]. Annak ellenére, hogy bizonyos citokróm P450 enzimek expressziója gyenge vagy hiányzik a normál hepatocitákhoz képest, a HepG2 metabolikus profilja ösztönözte a sejtvonal módosítására irányuló erőfeszítéseket a gyógyszer-metabolizmus jobb tanulmányozása érdekében [13]. Az MCF7, PC3, 143B és HEK293 tumorsejtvonalakhoz képest a HepG2 sejtek egyedülálló aminosav-tartalmi profilokat mutatnak, amelyek jelentősen befolyásolják a fehérjeszintézist és a szekréciót, kiemelve egyedi metabolikus útjaikat [14].
A májbetegségek kutatásának feltárása a HepG2 segítségével
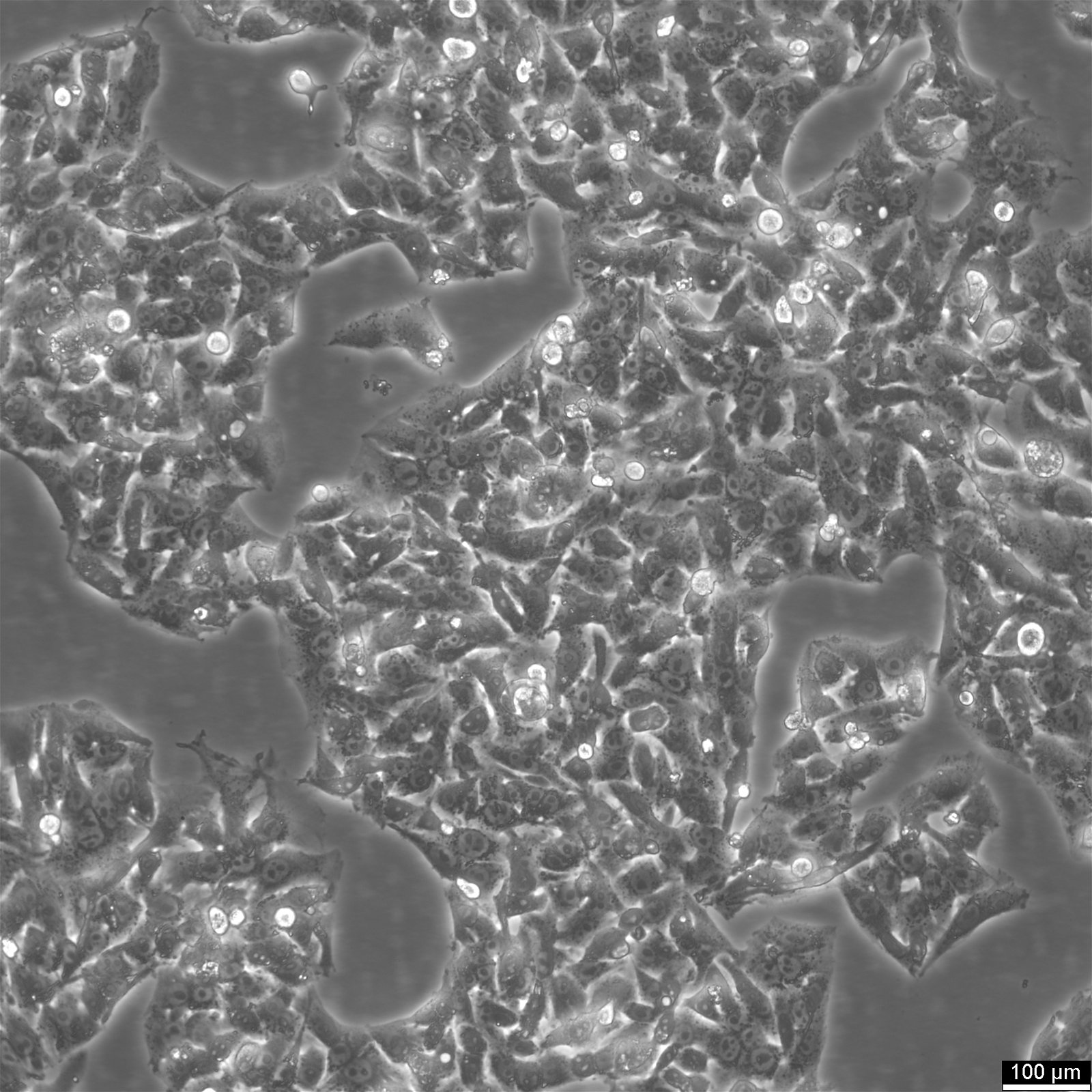
HepG2 sejtek átültetése
Az alábbiakban bemutatjuk az Accutase segítségével a tapadó sejtek sejtkultúra-lombikokból történő eltávolításának öt lépését:
- Távolítsa el a táptalajt a sejtkultúra-lombikból, és öblítse le az adhezív sejteket kalcium- és magnéziummentes PBS-sel. Használjon 3–5 ml PBS-t a T25-ös lombikokhoz és 5–10 ml-t a T75-ös lombikokhoz.
- Adjon Accutase-t a sejtkultúra-lombikba, 1–2 ml-t T25-ös és 2,5 ml-t T75-ös lombikokhoz. Győződjön meg arról, hogy az Accutase az egész sejtréteget befedi.
- Inkubálja a lombikot szobahőmérsékleten 8–10 percig.
- Óvatosan szuszpendálja újra a sejteket 10 ml friss táptalajjal.
- Centrifugálja az újra szuszpendált sejteket 5 percig 300xg-en, szuszpendálja őket friss táptalajban, majd adagolja őket friss táptalajt tartalmazó új lombikokba.
A HepG2 sejtek jövőbeli kilátásai
A HepG2 sejtvonal teljes potenciáljának feltárására irányuló kutatások folytatódnak, és áttörő eredményeket értek el a citokrómok expressziójának növelésében. A kutatók emellett a háromdimenziós szferoid sejtkultúrák lehetőségét is vizsgálják, amelyek fiziológiailag relevánsabb rendszert kínálnak. A citokrómokat is magában foglaló metabolikus aktivitás a 3D-s szferoid HepG2-modellekben jelentősen magasabb, mint a 2D-s sejtekben, ami közelebb visz minket egy olyan modell létrehozásához, amely tükrözi a normális hepatocitákat. Ezenkívül a sejtfelszíni fehérjék helytelen eloszlásának hátterében álló dinamikus folyamatok feltárása elősegítheti a májbetegségek jobb megértését.
HepG2 sejtek: szerepük és jellemzőik az orvostudományi kutatásban – GYIK
Hivatkozások
- Vyas, R.C., Darroudi, F., Natarajan, A.T. Sugárzás által kiváltott kromoszómális törés és újraegyesülés emberi limfociták interfázis-metafázis kromoszómáiban, Mutat Res, 1991; 249(1):29-35.
- Woodfield, S.E., Shi, Y., Patel, R.H., Chen, Z., Shah, A.P., Srivastava, R.K., Whitlock, R.S., Ibarra, A.M., Larson, S.R., Sarabia, S.F., et al. MDM4 gátlás: egy új terápiás stratégia a P53 reaktiválására hepatoblastómában. Sci. Rep. 2021, 11, 2967.
- Hussain, S.P., Schwank, J., Staib, F., Wang, X.W., Harris, C.C. TP53 mutációk és hepatocelluláris karcinóma: betekintés a májrák etiológiájába és patogenezisébe. Oncogene 2004.
- Schicht, G., Seidemann, L., Haensel, R., Seehofer, D., Damm, G. A HepG2 és Huh7 hepatoma sejtvonalak használhatóságának kritikai vizsgálata a reszekálható hepatocelluláris karcinóma metabolikus reprezentációjának modelljeként. Cancers 2022, 14(17), 4227.
- Verrier, E.R., Colpitts, C.C., Schuster, C., Zeisel, M.B., Baumert, T.F. Sejtkultúra-modellek a hepatitis B és D vírusfertőzés vizsgálatához. Viruses 2016, 8, 261.
- Verrier, E.R., Colpitts, C.C., Bach, C., Heydmann, L., Weiss, A., Renaud, M., Durand, S.C., Habersetzer, F., Durantel, D., AbouJaoudé, G., et al. A célzott funkcionális RNS-interferencia szűrés feltárja, hogy a glikopán 5 a hepatitis B és D vírusok behatolási tényezője. Hepatology 2016, 63, 35–48.
- Gripon, P., Rumin, S., Urban, S., Le Seyec, J., Glaise, D., Cannie, I., Guyomard, C., Lucas, J., Trepo, C., Guguen-Guillouzo, C. Emberi hepatoma sejtvonal fertőzése hepatitis B vírussal. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660.
- Mersch-Sundermann, V., Knasmüller, S., Wu, X.J., Darroudi, F., Kassie, F. Emberi eredetű májsejtvonal alkalmazása citoprotektív, antigénotoxikus és kogenotoxikus szerek kimutatására. Toxicology. 2004; 198(1–3): 329–340.
- Fanelli, A. HepG2 (májhepatocelluláris karcinóma): sejtkultúra. HepG2. Letöltve: 2017. december 3.
- Xuan, J., Chen, S., Ning, B., Tolleson, W.H., Guo, L. A citokróm P450-eket expresszáló, HepG2-ből származó sejtek kifejlesztése az anyagcserével összefüggő, gyógyszer által kiváltott májtoxicitás értékelésére. Physiol. Behav. 2017, 176, 139–148.
- Ooka, M., Lynch, C., Xia, M. Az in vitro metabolizmusaktiválás alkalmazása nagy áteresztőképességű szűrésben. Int. J. Mol. Sci. 2020, 21, 8182.
- Huang, L., Coughtrie, M.W.H., Hsu, H. A dehidroepiandroszteron-szulfotranszferáz gén leregulációja humán hepatocelluláris karcinómában. Mol. Cell. Endocrinol.
- Zhu, Z., Hao, X., Yan, M., et al. A rákos őssejtek/elősejtek nagy arányban vannak jelen a CD133 + CD44 + populációban a hepatocelluláris karcinómában. Int J Cancer. 2010; 126:2067-2078.
- Arbus, C., Benyamina, A., Llorca, P.-M., Baylé, F., Bromet, N., Massiere, F., Garay, R.P., Hameg, A. A ciamemazin metabolizmusában részt vevő humán citokróm P450 enzimek jellemzése. Eur J Pharm Sci. 2007 dec.; 32(4-5):357-66.